1.21: Метаболічна різноманітність
- Page ID
- 7092

Уроки від диваків
Іноді розуміння процесу допомагає дивлячись на варіації процесу. Ми вивчаємо, як організми задовольняють свої енергетичні та матеріальні потреби. Основним матеріалом, який ми зараз розглядаємо, є вуглець, але він, як правило, приносить з собою водень і кисень як молекулу вуглеводів. Ми дійдемо до інших матеріалів в наступному розділі, але вуглець, водень і кисень складають 95% більшості організмів. Спочатку ми розглянемо загальні закономірності, знайдені в більшості організмів. Потім ми розглянемо кілька груп організмів, енергія та обробка вуглецю яких ексцентричні різними способами.
ТЕМИ
- Огляд: «нормальні» закономірності, що включають речовину та енергію
- Придбання енергії — клітинне дихання
- Придбання харчових гетеротрофів та фотосинтетичних автотрофів
- Придбання вуглецево-гетеротрофів та фотосинтетичних автотрофів
- «Ексцентричні» організми
- Отримання енергії, коли немає кисню
- ферментатори
- за допомогою альтернативних акцепторів електронів
- Хемоавтотрофи, що роблять їжу (вуглеводи) без світла
- Фотоавтотрофи, що розділяють речовину і енергію
- Отримання енергії, коли немає кисню
«Норма» — найпоширеніші процеси
1. Придбання метаболічної енергії
Майже всі організми використовують реакції, розглянуті в розділі про клітинне дихання, для забезпечення своїх енергетичних потреб: клітинне дихання забезпечує енергію у вигляді АТФ до тих пір, поки є запас вуглеводів і кисню. Кисень легко доступний у всіх земних середовищах існування через те, що в атмосфері 20% кисню, а місцеві виснаження рідкісні, оскільки атмосфера постійно змішується вітрами. На відміну від цього, в деяких водних ситуаціях (включаючи ґрунти, насичені водою) кисню може бути дефіцит. У таких ситуаціях присутній ряд організмів, як прокаріотичних (бактерій або архей), так і еукаріотичних, які можуть отримувати енергію АТФ за відсутності кисню, таким чином, що використовує лише гліколізну частину клітинного дихання. Крім того, існують також організми, виключно прокаріоти, які мають механізми виготовлення АТФ, які взагалі не покладаються на шляхи клітинного дихання.
2. Виготовлення власної їжі: потік енергії в фотосинтетичних автотрофах
Якщо уважно розглянути це, енергетика фотосинтетичних організмів здається надмірно залученою: світлові реакції виробляють АТФ і НАДПГ для того, щоб синтезувати вуглеводи з вуглекислого газу. Потім вуглеводи розщеплюються в клітинному диханні, щоб виробляти NADH (для наших цілей такий же, як NADPH) і АТФ. Навіщо турбувати виготовлення вуглеводів; чому б просто не використовувати світлові реакції для отримання «енергетичної валюти», АТФ і NADH? В обмеженій мірі фотосинтезуючі організми DO задовольняють деякі свої енергетичні потреби безпосередньо поза світловими реакціями, тобто не всі вироблені АТФ і НАДФГ використовуються для виготовлення вуглеводів, деякі живлення інших процесів. Але здебільшого потік енергії в фотосинтетичних організмах йде від світла до АТФ і НАДПГ, до вуглеводів, а потім назад до АТФ і NADH; і кількість доступної енергії зменшується по дорозі, оскільки передача енергії не є 100% ефективною. Існує кілька обґрунтувань такої поведінки фотосинтетичних організмів:
- клітинне дихання розвинулося першим, і фотосинтез з'явився як засіб, що дозволяє організмам уникнути необхідності «просити і красти їжу»
- потреба в енергетичних сполуках є безперервною, але нічний час запобігає фотосинтезу. АТФ та НАДГ не можуть бути легко збережені, і, хоча вони постійно переробляються, абсолютна кількість ADP/ATP та NADP/NADPH невелика. Навпаки, вуглеводи можуть зберігатися і можуть бути присутніми в набагато більшій кількості.
- для судинних рослин нижньоземні частини потребують енергетичних сполук, але не можуть фотосинтезувати. Лабільна природа АТФ і НАДГ означає, що їх не можна транспортувати, тоді як вуглеводи можуть.
Але ми побачимо, що є деякі організми, які використовують світло в строго енергетичній ролі, не використовуючи його для виготовлення вуглеводів, які згодом використовуються для отримання енергії.
3. Матеріальні потреби, зокрема вуглець
Що стосується матеріальних потреб, здебільшого (або ви можете сказати «для більшості їхніх частин»!) , організми складаються з вуглеводів, або молекул, які виготовляються з вуглеводів, наприклад, амінокислоти білків, які в основному складаються з вуглеводів з азотом групи (ів), а іноді також сірчаної групи, або жирів, які виробляються з вуглеводів, які мали більшість їх кисню вилучено. Таким чином, будь-який організм повинен отримувати вуглеводи, щоб побудувати більше себе, тобто рости. Для фотосинтетичних автотрофів потреби у вуглеводах забезпечуються фотосинтезом, але зверніть увагу, що будь-які вуглеводи, спрямовані на матеріальні потреби, недоступні для використання в енергетичних потребах. Для гетеротрофів потреби у вуглеводах задовольняються шляхом привласнення частини того, що споживається на будь-які «будівельні проекти», які можуть вимагати гетеротроф, але, як наслідок, зменшуючи кількість енергії, яку можна отримати. Майже всі організми є або гетеротрофами, споживання органічного матеріалу (тіла або частини тіл організмів) забезпечує їх як енергією, так і вуглецевими матеріалами або фотосинтетичними автотрофами, які використовують світло, щоб зробити власні вуглеводи з вуглекислого газу і води, а потім, як гетеротрофи, використовують цю «їжу» як для матеріальних, так і для енергетичних потреб. Однак є деякі організми (всі прокаріотичні), які є нефотосинтетичними автотрофами, тобто здатні виробляти вуглеводи без сонячного світла, використовуючи хімічні речовини в якості енергопостачання. Існують також організми, знову ж таки прокаріоти, чиї харчові звички включають або вимагають деяких матеріалів, які явно не є «органічними», тобто виготовленими організмами, наприклад, формальдегідом або «пластиком» (поліетилентерефталат). Є також деякі організми, які «їдять» виключно для вуглецевого харчування і мають інші механізми, які не засновані на вуглеводній їжі, для отримання енергії.
Нижче наведені деякі групи ексцентричних організмів, організмів, які відрізняються від більш звичних закономірностей, розглянутих вище.
Ексцентричні організми I: анаеробні організми — використання альтернативних акцепторів електронів
Пам'ятайте, що роль кисню в клітинному диханні повинна бути «електронним магнітом», низькою точкою, до якої надходять електрони. Електрони, отримані з вуглеводів, переносяться в форму NAD + (утворюючи NADH) і з нього електрони протікають через ряд носіїв ланцюга транспорту електронів. Роль кисню має важливе значення як тому, що він «керує» потоком електронів, який, в свою чергу, дозволяє синтезувати АТФ, а також тому, що процес регенерує NAD +, який необхідний як для гліколізу, так і для циклу лимонної кислоти для продовження. У більшості земних середовищ існування кисню багато і з тих пір фотосинтез став популярним приблизно 2 мільярди років тому. Але бувають ситуації, коли кисень стає дефіцитним, як правило, в результаті клітинного дихання в поєднанні з фізичними факторами, наприклад, перезволоженими грунтами, які роблять поповнення кисню малоймовірним (Глава 26). Одне рішення дефіциту кисню полягає в тому, щоб знайти інший атом або молекулу, яка буде виконувати ту ж роль, тобто бути «ситуацією спуску», до якої можуть надходити електрони.
Є ряд бактерій, які роблять це, використовуючи ряд різних молекул як замінники кисню. Особливо важливою групою для харчування рослин називають денітрифікуючі бактерії. Вони використовують нітрат (NO 3 -) як акцептор електронів, який приймає електрони від NADH, що утворюються в гліколізі та циклі лимонної кислоти. У процесі вони перетворюють нітрат, форму азоту, яку більшість рослин можуть засвоювати (тобто утилізувати) в азотний газ (N 2), форму азоту, яку рослини не можуть асимілювати (хоча деякі можуть за допомогою симбіонта). Більш того, N 2 летючий і може виходити з грунту; на відміну від цього, нітрат є іоном і, отже, не в змозі покинути ґрунтовий розчин, хоча сам ґрунтовий розчин може покинути грунт, забираючи з собою іони (тобто вилуговування). Процес перетворення нітрату в азотний газ називається денітрифікацією, а в деяких ситуаціях він спричиняє значні втрати азоту з ґрунтів. Ще одним замінником кисню є сульфат (SO 4 2-), який після прийняття електронів перетворюється в сірководень. Як і у випадку з денітрифікацією, ці реакції негативно впливають на мінеральне живлення рослин, усуваючи форму, яку рослина може засвоїти (сульфат), і вводячи сірку в форму, яку рослини менш здатні придбати, а також форму, яка є летючою і може бути втрачена з місця проживання рослини.
Ексцентричні організми II: анаеробні організми - ферментація
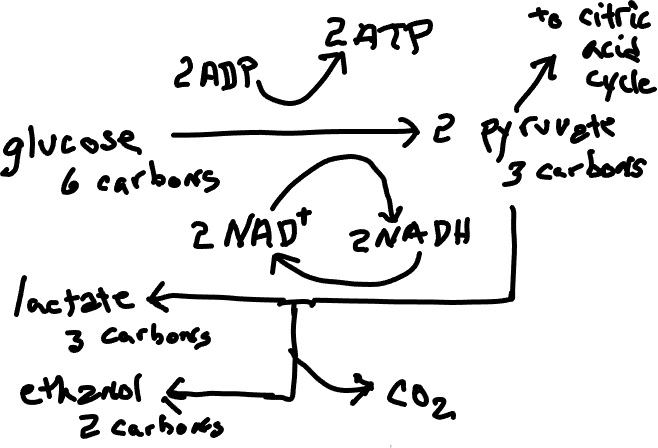
Одним з рішень дефіциту кисню, і найбільш близьким до нормальних моделей, є процес, який називається ферментацією («анаеробне дихання»), який включає додаток до гліколізу, першу стадію клітинного дихання і обов'язково усунення решти частин клітинного дихання. Піруват, що виробляється гліколізом, не проходить цикл лимонної кислоти, а замість цього використовується прямо або опосередковано для прийому електронів від NADH, дозволяючи регенерувати NAD + (рис. 1), що має важливе значення для гліколізу. Значним для ферментуючого організму є те, що це дозволяє протікати гліколізу, хоча в процесі отримують набагато менше АТФ, ніж було б утворено, якби цикл лимонної кислоти та окислювальне фосфорилювання змогли відбутися. Також важливим є те, що, хоча кінцевими продуктами клітинного дихання є вода і вуглекислий газ, доброякісні речовини, які легко диспергуються, кінцеві продукти бродіння, як правило, етанол або молочна кислота, більш токсичні і важкі для усунення. Ферментація надзвичайно важлива в людських справах, як при виробництві алкогольних напоїв (як правило, грибками, але зрідка бактеріями), так і при виробництві бажаних харчових продуктів, наприклад, закваски (рис. 2), квашеної капусти, сметани, йогурту, на всі які впливає вироблення молочної кислоти. Деякі ферментуючі організми є факультативними анаероби (тобто вони можуть жити з киснем або без нього). Більшість з них переходять на бродіння, якщо кисню немає, але деякі здійснюють бродіння незалежно від наявності кисню. Деякі з них є облігатними анаероби, що означає, що вони не можуть жити в присутності кисню.

Ексцентричні організми III: приготування їжі без сонячного світла - хемосинтетичні організми
Більшість автотрофів («самоїдів») роблять їжу за допомогою фотосинтезу, а потім їдять самі. Енергія світла - це те, що робить несприятливу реакцію, зменшення вуглекислого газу водою, більш імовірним. Розпливчастим, але загальним описом процесу є те, що частина енергії світла захоплюється рослиною і зберігається у вигляді вуглеводів. Більш конкретний опис полягає в тому, що світло здатне в організованій структурі мембрани переміщати електрони таким чином, що NADPH утворюється з NADP + за допомогою електронів, отриманих з води. В процесі АТФ утворюється в результаті потоку електронів. У циклі Кальвіна ці продукти, NADPH та АТФ, можуть спричинити зменшення вуглецю електронами, наданими NADPH, та АТФ, що сприяє реакціям. Світло має важливе значення для фотосинтезу, оскільки забезпечує механізм отримання NADPH та АТФ. Але ці метаболіти можуть бути зроблені іншими способами, і є нефотосинтезуючі автотрофи, які роблять саме це. Вони описуються як хемосинтезирующие організми, і вони виробляють власну їжу способами, які не потребують світла. Потрібна хімічна речовина, яка може пожертвувати електрони для зменшення NADP + до NADPH та мембранна система, яка дозволяє синтезувати АТФ, коли електрони надходять від донора до NADP +. Хемосинтетичні організми є рідкістю і зустрічаються лише всередині бактерій та архей, але вони можуть бути дуже значними в певних середовищах існування та при здійсненні процесів, важливих для глобальних біогеохімічних циклів. З точки зору рослин, найважливішою групою цих організмів є нітрифікуючі бактерії, група, яка окислює аміак (NH 3) до нітриту (NO 2 -), і група, яка окислює нітрит до нітратів (NO 3 -). Результатом двох реакцій є те, що аміак перетворюється на нітрат, який є кращим джерелом азоту для більшості рослин, і, на відміну від аміаку, не може вийти з ґрунту як газ. Зверніть увагу, що обидві ці реакції є окисленнями (це очевидно в реакції NH 3 до NO 2 - як водні речовини видаляються з азоту, але також вірно в NO 2 - до NO 3 - перетворення). Електрони, видалені з азотної сполуки, використовуються для зниження NADP + до NADPH і забезпечують потік електронів (через мембрану), що дозволяє синтезувати АТФ. Ці реагенти потім використовуються для зменшення вуглекислого газу в реакціях циклу Кальвіна, «фіксуючи» його у вуглеводній формі. Нітрифікуючі бактерії, як і фотосинтезуючі рослини, виробляють власну їжу, а потім використовують її для синтетичних реакцій (як будівельний матеріал) або як джерело енергії (оскільки він окислюється до вуглекислого газу в клітинному диханні). Існують інші хемосинтетичні організми, крім нітрифікуючих бактерій, включаючи бактерії в глибоководних вентиляційних отворах, які використовують сірководень як джерело енергії для фіксації вуглецю, метаногенів, які використовують енергію диводневого газу (H 2) для фіксації вуглецю (і в той же час виробляють метан) та метану «їдці», які окислюють метан до азоту (N 2), одночасно зменшуючи вуглець. Багато, але не всі, хемосинтезирующие організми є архебактеріями, хоча нітрифікуючі бактерії не є.
Ексцентричні організми IV: розділення речовини та енергії

Для більшості організмів «їжа», тобто те, що гетеротрофи поглинають або ковтають, а для фотосинтезаторів вуглеводи, які вони виробляють, відіграють подвійну роль: як джерело енергії (виробляє АТФ та НАДГ) та як джерело вуглецю, забезпечуючи «відновлені вуглеці», які використовуються в різних біосинтетичних реакціях, які в кінцевому підсумку може зробити організм більшим. Шляхи для двох процесів спочатку однакові, але якщо вуглець буде використовуватися як будівельний матеріал, виходить набагато менше енергії і утворюється менше вуглекислого газу. Якщо використовується для енергії, «їжа», тобто вуглеводи, закінчується як вуглекислий газ, а кількість отриманої енергії максимальна; якщо використовувати його для матеріалу, вуглеводи потрапляють у будь-яку з тисяч біомолекул, знайдених в організмі, в жирах, білках, нуклеїнових кислотах тощо, а також кількість енергії отриманий зменшується порівняно з тим, що станеться, якби всі вуглеці були повністю окислені до CO 2. Однак є деякі організми, які мають чіткі шляхи для отримання енергії, шляхи, які, як правило, взагалі не включають вуглець, і їх «харчування» полягає виключно для отримання атомів вуглецю для біосинтезу. Найпростіша група для розуміння - це фотогетеротрофи. Вони включають як археї, так і бактерії, які здатні використовувати сонячне світло в процесі, який дозволяє їм синтезувати АТФАле не таким чином, що виробляє вуглеводи.Таким чином, вони повинні «їсти їжу» (поглинати вуглеводи/органічні молекули) НЕ обов'язково для своїх енергетичних потреб (тобто для постачання АТФ), а скоріше для задовольняють свої потреби у вуглеці. Цей спосіб життя зустрічається у кількох археях, наприклад, Halobacterium, який має пігмент, бактеріородопсин, пов'язаний з родопсином, що міститься в очах хребетних. Бактеріородопсин - це мембранно-охоплюючий білок, який може набувати протони в цитозолі, змінювати конформацію за рахунок поглинання світла та вивільняти протони на зовнішній стороні мембрани (рис. 3). Потім протони потрапляють у клітину, вниз по їх електрохімічному градієнту, і, як це буває у світлових реакціях фотосинтезу та окислювального фосфорилювання в клітинному диханні, рух протонів викликає синтез АТФ з АДФ та неорганічного фосфату. Більшість фотогетеротрофів - це бактерії, які використовують форму хлорофілу для поглинання світлової енергії і знову створюють протонний градієнт, який можна використовувати для синтезу АТФ, але для матеріальних потреб потрібен джерело відновленого вуглецю.
Подібним чином існують хемогетеротрофи, організми, які «їдять» за свій матеріал (тобто зменшені потреби у вуглеці), але генерують АТФ хімічними засобами, які не включають клітинне дихання та окислення вуглеводів.
Таблиця 1 нижче узагальнює різні режими придбання речовини (вуглецю) та енергії (зменшення потужності та АТФ).
Таблиця 1.
|
енергія |
джерело вуглецю |
груп |
|
|
гетеротроф |
енергії (відновлювальної потужності і АТФ) від окислення зібраних біомолекул |
вуглець (відновлений) із зібраних біомолекул |
всі тварини, всі гриби, слиз цвілі, форми для води, деякі дінофлагелати, більшість бактерій, більшість археї |
|
фотосинтетичний автотроф |
енергії (відновлювальної потужності і АТФ) від окислення саморобних біомолекул |
вуглецю з вуглекислого газу відновлюється в фотосинтезі і використовується для синтезу біомолекул |
всі рослини, зелені водорості, червоні водорості, бурі водорості, діатомові водорості, деякі динофлагелати, криптомонади, ціанобактерії, зелені сірчані бактерії |
|
хемосинтетичний автотроф |
енергії (відновлювальної потужності і АТФ) від окислення саморобних біомолекул |
вуглецю з вуглекислого газу, відновленого у фотосинтезі, що використовується для синтезу біомолекул |
деякі, але не багато, бактерії (нітрифікуючі бактерії, сірчаноокислювальні бактерії, залізоокислювальні бактерії, деякі метаногени) і деякі архей (метаногени). |
|
фотогетеротроф |
енергія (деяке зниження потужності, але в основному АТФ) від реакцій, керованих світлом |
вуглець (відновлений) із зібраних біомолекул |
Деякі, але не багато, археї (Halobacterium), деякі зелені несірчані бактерії, деякі фіолетові несірчані бактерії. |
|
хемосинтетичний гетеротроф |
енергія (відновна потужність і АТФ) від електронного потоку, керованого неорганічними джерелами електронів |
вуглець (відновлений) із зібраних біомолекул |
деякі, але не багато, бактерії (Beggiatoa) і деякі архей (метаногени). |