1.20: Фотосинтез
- Page ID
- 7013

Рослини ростуть з повітря. У той час як невелика частина їх (сухої маси) маси надходить з грунту, приблизно 98% надходить з атмосфери. Типова порція картоплі - 6 унцій = 170 грам. Оскільки картопля становить приблизно 80% води, порція картоплі становить близько 34 грамів сухої ваги. Ця маса являє собою майже чистий крохмаль, який має хімічну формулу (C 6 H 12 O 6). Атоми вуглецю становлять 40% від ваги крохмалю (або будь-якого гексози або гексози полімеру), так що 34 грами картоплі мають 13,6 грама вуглецю = 1,13 моль вуглецю. Це було отримано з повітря, яке становить приблизно 400 проміле вуглекислого газу. Для того, щоб придбати вуглець, необхідний для порції картоплі, рослині доводиться видобувати вуглець приблизно з 67 тисяч літрів повітря. Ця фотосинтетична здатність ще більш примітна, оскільки передбачає передачу енергії світла (електромагнітного випромінювання) в хімічну енергію, присутню у вуглеводі, який виробляється. Синтезовані гексози мають значно більше енергії, ніж сировина (вуглекислий газ і вода), що використовується для їх виробництва.
ТЕМИ
- Огляд
- Світлозалежні реакції
- Реакції засвоєння вуглекислого газу
- Фотосинтетичні обмеження: компроміс фотосинтезу/транспірації
- C4 фотосинтез
- Фотосинтез CAM
Огляд
Фотосинтез виробляє вуглеводи. Назва показує, що вони спочатку вважалися: гідратовані вуглеці (тобто вода, додана до вуглецю), але дослідження показали, що більш точний опис вуглеводів - це зменшений вуглекислий газ. Це проілюстровано в загальному рівнянні фотосинтезу:
СО 2 + Н 2 О —> (СН 2 О) п + О 2
До вуглекислого газу додається водень, тобто він відновлюється, а з кисню видаляється водень, тобто окислюється. Зауважимо, що, як і у випадку з клітинним диханням, загальне рівняння є резюме чистого ефекту декількох реакцій, що відбуваються одночасно. Зокрема, при фотосинтезі вуглекислий газ НЕ реагує з водою. Натомість і вода, і вуглекислий газ споживаються в групі реакцій, які в кінцевому підсумку виробляють кисень і вуглеводи.
Загальна реакція, як написано, є винятковою, оскільки кисень є високоелектронегативним атомом, який сильно притягує водні гени, набагато сильніше, ніж атом вуглецю, до якого переносяться водні. З цієї причини реакція «піднімається» і навряд чи відбудеться, тоді як реакція в зворотному напрямку набагато частіше відбувається, при цьому вуглеводи окислюються киснем з утворенням вуглекислого газу та води (див. Попередній розділ). Останній розділ проілюстрував, що клітинне дихання (окислення вуглеводів) «кероване» електронегативністю кисню; ця електронегативність «тягне» електрони через внутрішню мітохондріальну мембрану, в кінцевому підсумку об'єднуючи їх з киснем. Таким чином, важливим питанням щодо фотосинтезу є те, що витягує електрони від кисню молекули води - що має сильніший «тяга» для водню, ніж кисень (тобто, що є більш потужним окислювачем, ніж кисень)? Світло, взаємодіючи з пігментом хлорофілом, відіграє вирішальну роль у створенні сильного окислювача. Також критичним є структура хлоропласту, який, як і мітохондрії, складається зі складної структури мембран, що розділяють водні відсіки. Мембрани містять хлорофіл (рис. 1), а також інші пігменти і білки, організовані дуже специфічними способами.

Світлозалежні реакції фотосинтезу
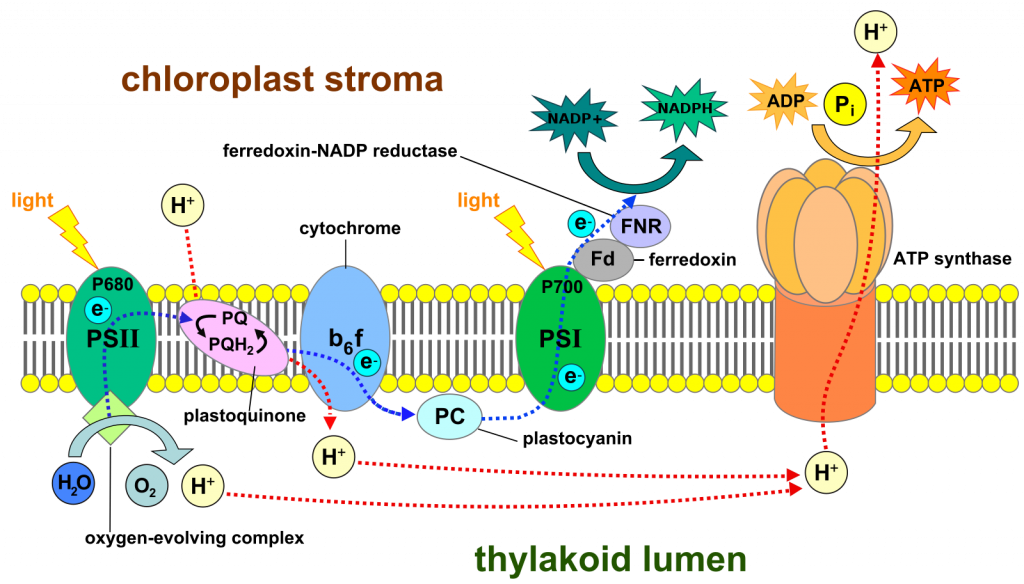
Роль світла у фотосинтезі полягає в перестановці конкретних молекул хлорофілу, змушуючи їх втрачати електрони (окислюючи їх) і тим самим роблячи їх окислювачами, які видаляють електрони з інших молекул і в кінцевому підсумку утворюють воду.Світло є формою енергії і, отже, здатний змінювати обставини матеріал, з яким він взаємодіє. Зокрема, світло змінює електронну конфігурацію хлорофілу, зміщуючи електрон зі свого нормального положення в ситуацію, коли він з більшою ймовірністю вирветься з молекули хлорофілу, тобто після поглинання світла хлорофіл швидше окислюється. Окислення хлорофілу робиться ще більш імовірним, оскільки поблизу знаходиться молекула у внутрішній мембрані хлоропласту, яка здатна приймати електрон. Електрон, втрачений від хлорофілу, в кінцевому підсумку (після багатьох кроків) закінчується пов'язаним з вуглецем молекули вуглекислого газу, утворюючи вуглеводи. Окислена молекула хлорофілу (тобто та, яка відсутня електрон) не є достатньо сильним окислювачем для вилучення електронів з води. Але він може діяти на «комплекс, що розвивається кисню», ферментний комплекс, який містить чотири атоми марганцю. Хлорофіл, який не має електрона, здатний окислювати один з чотирьох атомів марганцю. Після того, як цей процес повторюється ще три рази і всі чотири атоми марганцю окислюються, молекула тепер є досить сильним окислювачем, щоб діяти на дві молекули води, видаляючи чотири електрони, по одному з чотирьох воднів, і виробляючи чотири протони (H +) і одну молекулу O 2.
Електрон, втрачений від хлорофілу, йде шляхом через мембрану, подібну потоку електронів через внутрішню мітохондріальну мембрану в процесі «електронного транспорту» (= окислювального фосфорилювання) клітинного дихання, іноді використовуючи подібні носії електронів.
І, подібно до деяких етапів окислювального фосфорилювання, деякі перенесення електронів мають ефект переміщення протонів з одного боку мембрани на іншу. Більше того, «розщеплення» води, що виконується марганцевмісним білком, додає протони на ту саму сторону мембрани. Накопичення протонів на одній стороні мембрани створює електрохімічний градієнт поперек мембрани. І, через електрохімічного градієнта, АТФ може синтезуватися таким, яким він є в електронно-транспортному ланцюзі клітинного дихання. Але в цьому випадку електрони не надходять до кисню. Вони рухаються спочатку до іншого атома хлорофілу, але тільки після того, як він, як описана раніше молекула хлорофілу, окислюється під дією світла. Коли цей другий хлорофіл отримує електрон з ланцюга транспорту електронів, він перетворюється назад в нормальний стан, і в цьому стані хлорофіл може знову поглинати світло, збуджуватися і втрачати електрон, продовжуючи таким чином процес. Зверніть увагу, що ми виділили дві різні молекули хлорофілу, обидві з яких поглинають світло і втрачають електрони. Одна молекула хлорофілу (звана хлорофілом 680) отримує «замінні» електрони з білка, що містить марганець; другий хлорофіл (званий хлорофілом 700) отримує електрони заміщення з ланцюга транспорту електронів. Потік електронів, описаний до цих пір, становить:
вода—>
ферментний комплекс марганцю —>
хлорофіл 680—>
ланцюг транспорту електронів —>
хлорофіл 700.

Але вони ще не закінчили свій шлях! Зрештою, ці електрони будуть відновлювати вуглекислий газ, але перед тим, як потрапити до вуглецю вуглекислого газу, вони переносяться на інший важливий проміжний продукт, NADP + (рис. 2), сполуку, дуже схожу на NAD +, яка діє в мітохондріях під час клітинного дихання. Як і NAD +, NADP + може приймати два електрони та протон для утворення NADPH і може втратити ті ж елементи для реформування NADP +, тобто це носій електронів, який (як NADP +) може окислювати сполуки і (як NADPH) може зменшувати сполуки. NADP+ отримує електрони від молекул носіїв, які отримують їх від «збудженої» молекули хлорофілу 700. NADPHє відносно стабільною молекулою і водорозчинний, на відміну від багатьох електронних носіїв, що беруть участь у фотосинтезі, які розчинні тільки в ліпідах мембрани хлоропласту.
Загалом, те, що ми описали до цих пір, - це світловий потік електронів від води до NADP +, утворюючи NADPH і O 2 (рис. 3). Потік відбувається у внутрішній мембрані хлоропласту і включає два етапи, де енергія світла є значною для того, щоб зробити перенесення електронів (тобто окислювально-відновні реакції) більш імовірним. Потік електронів через мембрану здатний створити протонний градієнт, як це робить у внутрішній мембрані мітохондрій. І, як і у випадку з мітохондріями, цей градієнт у протонів може бути використаний для синтезу АТФ. Ця група реакцій, що працюють від світла і створюють NADPH та АТФ з NADP +, ADP та неорганічного фосфату, називається «світловими реакціями» і підсумовується як:
Н 2 О + НАДП + + АДП + ІП —> НАДФ + АТП + О 2
Реакції асиміляції вуглекислого газу фотосинтезу
Продукти світлових реакцій, NADPH і АТФ, використовуються для синтезу вуглеводів з вуглекислого газу, процес, який називається фіксацією вуглекислого газу.
Фіксація вуглецю принципово передбачає використання «відновної потужності» NADPH для зменшення вуглекислого газу, і процес узагальнений у наступному рівнянні:
НАДФ+АТП + СО2—> (СН 2 О Н) + НАДП + АДП + Пі
Зауважимо, що одним з надзвичайно важливих аспектів цих реакцій є те, що вона регенерує метаболіти, необхідні в світлових реакціях: NADP +, ADP та iP. Оскільки запаси цих метаболітів обмежені, дуже важливо, щоб вони були перероблені. Реакції фіксації вуглецю можна узагальнити в три етапи: карбоксилювання, відновлення та регенерація.
карбоксилювання
Карбоксилювання описує включення вуглекислого газу в органічну молекулу. Цікаво, що це може бути досягнуто без залучення будь-якого з продуктів світлової реакції. Карбоксилювання відбувається, коли вуглекислий газ додається до метаболіту, який називається рибулоза бісфосфат (RubP), п'ятивуглецевий цукор з двома фосфатами, в реакції, каталізованої ферментом, який називається рибулоза бісфосфаткарбоксилаза (рубіско). Отримане 6-вуглецеве з'єднання швидко розпадається до двох молекул тривуглецевого з'єднання, званого фосфогліцерином (PGA).
Зменшення
Хоча вуглекислий газ був засвоєний, PGA не є дуже корисною сполукою, оскільки вона занадто окислена. Щоб бути корисним, PGA потрібно зменшити. Потім його можна використовувати як молекулу-попередник для створення різноманітних біомолекул, таких як цукру, амінокислоти, нуклеїнові кислоти та багато інших. Крім того, відновлену сполуку можна використовувати для отримання більшої кількості РУБП і, таким чином, дозволити засвоювати більше вуглекислого газу.
Зниження PGA здійснюється за допомогою NADPH і АТФ, що утворюються в світлових реакціях фотосинтезу, і виробляє тривуглецевий цукор, який називається гліцеральдегід-3-фосфат (G3P).
ПАГ+НАДФ+АТП —> 3П+НАП + АДП + Пі
Регенерація рубП
Для підтримки фотосинтезу рослині необхідно регенерувати RUBP, 5-вуглецевий цукор, який використовується для придбання CO 2. Це відбувається при синтезі RUBP з G3P. Очевидно, що ви не можете зробити п'ятивуглецевий цукор з трьох вуглецевих цукру. Ви можете зробити це за допомогою двох молекул G3P, але залишився б один «фіксований» вуглець. Однак синтез може бути здійснений акуратно, якщо почати з п'яти молекул G3P (п'ятнадцять загальних вуглеців) і робить три рублі (також 15 вуглецю). Ці реакції називаються циклом Кальвіна-Бенсона, і вони вимагають одного АТФ, зробленого в легких реакціях для кожного виробленого РУБП.
У той же час G3P можна використовувати для виготовлення шестивуглецевих цукрів, зокрема глюкози та фруктози, а з них сахарози, крохмалю, целюлози та найрізноманітніших полісахаридів.
Поклавши обидві ці дії разом, якщо шість молекул вуглекислого газу фіксуються карбоксилюванням шести RUBP, то 12 G3P може бути отриманий після відновлення з використанням 12 NADPH і 12 АТФ. Десять молекул G3P можуть бути використані для регенерації шести рублів, і цей процес вимагає ще шести АТФ. Решта дві молекули G3P можуть бути використані для утворення фруктози або глюкози. Саме так всі рослини здійснюють фотосинтез. Для кожного асимільованого вуглекислого газу потрібно два НАДПГ і три АТФ.
Зверніть увагу, що ВСІ перераховані вище рівняння є резюме декількох реакцій, що відбуваються одночасно. Існує велика кількість хімії, що відбувається в хлоропластах, хоча чистий ефект може бути виражений просто як
6 СО 2 + 6 Н 2 О —> (С 6 Н 1 2 0 6) + 6 О 2
Зауважимо, що до зрівняння не входять АТФ, ADP, IP, NADP +, NADPH, RUBP, PGA або G3P. Всі ці реагенти виробляються з тією ж швидкістю, що і витрачаються. Чисте рівняння також приховує той факт, що насправді 12 Н 2 О потрібні як реагенти і що 6 Н 2 О є продуктами (чистий ефект - це просто витрата 6 Н 2 О). Частина елегантності фотосинтезу полягає в тому, що, незважаючи на безліч реакцій, що відбуваються, чистий ефект дуже простий. Ще один аспект його елегантності полягає в тому, що продукт процесу, вуглевод (CH 2 O) n, може бути використаний для виготовлення всіх різноманітних хімічних речовин, які виробляє рослина, не тільки очевидні (крохмаль, целюлоза, геміцелюлоза, пектини), але менш очевидні (амінокислоти, нуклеїнові кислоти , ліпіди, лігнін та ін.). Більше того, вуглеводи також використовуються як джерело енергії для живлення цих синтетичних реакцій: У клітинному диханні. Вуглевод споживається для отримання вуглекислого газу та води під час виробленняATP та NADH, хімічних речовин, необхідних у багатьох синтетичних реакціях.
Придбання вуглекислого газу, проблеми та рішення
Вуглекислий газ, зафіксований у фотосинтезі, виводиться з атмосфери. За останні 150 років рівень вуглекислого газу в атмосфері збільшився більш ніж на 40%, але вони все ще дуже, дуже низькі - 0,04% або 400 ppm (частин на мільйон; на кожен мільйон молекул в повітрі лише 400 з них є вуглекислим газом). Вуглекислий газ потрапляє в лист шляхом дифузії, але оскільки максимальна концентрація зовні становить 400 проміле, а для більшості рослин мінімальна концентрація всередині становить близько 100 проміле. Цей мінімум є результатом того, що rubisco не може карбоксилювати RUBP, якщо концентрація вуглекислого газу нижче 100 ppm, отже, [CO 2] не може йти нижче 100 ppm. Отже, рушійна сила дифузії (різниця в [СО 2] між внутрішньою і зовнішньою стороною листа) низька і потенційно обмежує потік вуглекислого газу в лист і, отже, фотосинтез. З огляду на рівняння потоку (глава 3), потік вуглекислого газу в лист можна збільшити за рахунок зменшення опору руху, тобто, зробивши лист більш пористим до атмосфери шляхом відкриття стоматів, регульованих пор в епідермісі листя. На жаль для більшості рослин цей вплив атмосфери призводить до втрати води, що називається транспірацією. Втрачену воду потрібно замінити водою з грунту, водою, якої іноді не вистачає. Це призводить до того, що було названо компромісом фотосинтезу/транспірації: настільки, наскільки рослина отримує вуглекислий газ для фотосинтезу, вона втрачає воду при транспірації.
Низькі концентрації вуглекислого газу в повітрі посилюють проблему: дозволити більшій кількості вуглекислого газу потрапляти в лист може більш повно відкрити його стомати, але це призводить до втрати більшої кількості води. У вологих середовищах, де вода легко доступна, немає проблем з дуже пористими листям (тобто відкритими стоматами), але коли води дефіцит, рослина повинна збалансувати прирост вуглецю з втратою води. Стоматальна поведінка відображає цей компроміс: вони, як правило, закриваються вночі, коли фотосинтез неможливий, а під час посухи, при придбанні води замінити втрачену важко.
Додатковий вуглекислий газ в атмосфері за останні 200 років, ймовірно, пояснює спостереження, що земля «зеленіша» (збільшений покрив листя) зараз, ніж 50 років тому. Більше вуглекислого газу може діяти як «добриво» та/або дозволяє рослинам вижити в умовах низького водопостачання.
Хоча існує ряд особливостей, які дозволяють деяким рослинам виживати в сухих умовахіснує така, яка безпосередньо передбачає розглянуті вище фотосинтетичні реакції. Цей модифікований фотосинтетичний шлях, який називається шляхом С4, дозволяє деяким рослинам купувати вуглекислий газ, втрачаючи менше води, ніж звичайна рослина. Рослини С4 концентрують вуглекислий газ у відносно невеликій частині листа, званої оболонкою пучка клітин. Шляхи утилізують дві карбоксиляції.Перший відбувається в клітині мезофілу листя і використовує фермент, ПЕП-карбоксилазу, який додає вуглець до трьох вуглецевих сполук, фосфоенолпіруват (PEP). Карбоксилаза PEP може працювати при рівнях вуглекислого газу до приблизно 10 проміле, приблизно 1/10 від концентрації, необхідної для роботи руббіско. Чотири вуглецеві сполуки, що виробляються карбоксилазою ПЕП, транспортується через плазмодезмати до оболонки збільшених клітин (рис. 4), яка оточує судинні нитки листа. Тут чотиривуглецеве з'єднання декарбоксилюється, виділяючи вуглекислий газ, який згодом фіксується в нормальному фотосинтезі за допомогою руббіско. Решта три вуглецевих фрагмента транспортуються назад в клітини мезофілу. Використання цієї системи двох карбоксиляцій дозволяє рослинам виробляти середовище (клітини оболонки пучка), де є більш висока концентрація вуглекислого газу. Тільки в цьому місці, де присутній рубіско. Через механізм концентрації CO 2, [CO 2] може бути більше 100 ppm в комірках оболонки пучка, тоді як [CO 2] повітря всередині листа близько до менше 10 ppm. Оскільки концентрація CO 2 істотно нижча, ніж у звичайному листі, може бути більша рушійна сила дифузії в лист. Це дозволяє опір бути вищим (стомати більш закриті), при цьому досягаючи такої ж кількості потоку вуглекислого газу (фотосинтез), як рослина, яка не використовувала цей шлях. Таким чином, лист може функціонувати фотосинтетично, будучи набагато менш пористим до вуглекислого газу, тим самим втрачаючи менше води при транспірації.

Рослини, які використовують цей подвійний шлях карбоксилювання, називаються «рослинами С4», оскільки перше карбоксилювання виробляє чотири вуглецеві сполуки, на відміну від моделі у більшості рослин, які називаються рослинами С3, де карбоксилювання виробляє три вуглецеві сполуки. Шляхи C4 викладено нижче:
- в мезофільних клітині свинцю три вуглецевої сполуки, ПЕП, карбоксилюється ПЕП-карбоксилазою, утворюючи чотири вуглецевої сполуки
- чотири вуглецеві з'єднання транспортується в клітини оболонки пучка, які оточують судинні пучки (жилки листа)
- чотири вуглецеві сполуки декарбоксильовані, виділяючи СО 2 і піруват, три вуглецеві сполуки
- піруват транспортується назад до клітин мезофілу, де він перетворюється в ПЕП в процесі, який вимагає енергії у вигляді НАДПГ і АТФ
- CO 2, який був випущений у оболонці пучка, фіксується, використовуючи рубіско в нормальному шляху
Зверніть увагу, що C4 вимагає всіх механізмів і реакцій фотосинтезу С3, це просто, що існує додатковий набір кроків перед малюнком C3. Фотосинтез С4 менш ефективний, ніж фотосинтез С3, оскільки він вимагає більше енергії АТФ. Однак він є більш ефективним з точки зору використання води, і це має більше значення в більш сухих регіонах. Крім того, з причин, в які ми не будемо вдаватися, фотосинтез C4 сприятливий при більш високих температурах.

Існує друга група рослин, які використовують подвійний шлях карбоксилювання, але модифікованим способом. Їх називають заводами CAM. CAM відноситься до метаболізму крассулацевої кислоти, оскільки модель поведінки, показана цією групою, була вперше виявлена у сукулентних рослин сімейства Crassulaceae. У рослин CAM є кілька особливостей: вони, як правило, соковиті, тобто мають товсті м'ясисті листя (рис. 5) або без листя і товстий м'ясистий стебло (наприклад, кактус). Вони, як правило, демонструють виражену добову картину кислотності тканин з найвищою кислотністю на світанку та зменшенням кислотності в денний час доби та підвищенням кислотності в нічний час. Найбільш своєрідним є те, що вони відкривають свої стомати вночі, а не вдень, тоді як більшість рослин відкривають свої стомати лише вдень, коли вони можуть фотосинтезуватися.
В основному ці рослини працюють так само, як це роблять рослини C4, але замість того, щоб мати просторовий поділ двох карбоксиляцій, як це мають рослини С4 (мезофіл проти оболонки пучка), вони мають тимчасове відділення (денний проти нічного часу). Початкове карбоксилювання відбувається вночі, коли стомати відкриті. Чотири вуглецеві сполуки є кислотою і викликає підвищення кислотності тканин. Протягом дня стомати закриваються, вуглекислий газ забезпечується декарбоксилювання чотирьох вуглецевих кислот і кислотність тканин знижується. Вивільнений вуглекислий газ повторно фіксується через рубіско з утворенням вуглеводів.
Чи фотосинтезуються ці рослини вночі або вдень? Це залежить від того, як можна визначити фотосинтез: придбання вуглецю відбувається вночі, але синтез цукру відбувається протягом дня. Фотосинтез CAM пов'язаний, але не обов'язково, з соковитою. Рослини CAM, як правило, зустрічаються в сухих середовищах існування, або кліматично, наприклад, пустель, або через мікросередовище існування, наприклад, епіфіти, рослини, які не вкорінені в землі, але ростуть на інших рослинях. Е піфіти часто піддаються посухи через відсутність зв'язку з грунтом. Кактус та інші безлисті сукуленти зазвичай є рослинами CAM. Швидкість фотосинтезу CAM дуже низькі, а темпи росту рослин CAM також низькі. Асоціація соковитості та CAM, ймовірно, відображає той факт, що рослини в посушливих середовищах існування часто розвивають соковитість для зберігання води і що сукулентність є більш сприятливим для CAM, оскільки соковиті тканини можуть зберігати більше вуглецю, як чотиривуглецева кислота.
Шаблони фотосинтезу CAM і C4 показують, що ці шляхи еволюціонували кілька разів, тобто шляхи мало говорять про філогенію. C4, C3 та CAM змішуються в пологах, сім'ях та порядках. Мабуть, це відносно «легко» для C4 та CAM розвиватися.