1.15: Пол і розмноження у насіннєвих рослин
- Page ID
- 7159

Насіння - чудове нововведення, яке було надзвичайно важливим для еволюції рослинного життя. Переважна більшість рослин, які ми спостерігаємо та використовуємо, мають насіння, а насіннєві рослини домінують у більшості наземних середовищ існування. Останній розділ описував, в загальних рисах, що таке насіння і які модифікації в загальному життєвому циклі рослин повинні були відбуватися чергування поколінь для того, щоб з'явилося насіння. Цей розділ заповнює деякі деталі для п'яти груп насіннєвих рослин:
- квітучі рослини, що мають 250 000 видів, на сьогоднішній день найрізноманітніша і повсюдна група рослин
- хвойні дерева: хоча мають лише 1000 видів, вони дуже часто зустрічаються та екологічно та економічно важливі
- цикади: невелика група близько 300 видів з обмеженим поширенням та значенням
- гнітофіти: невелика група, що складається лише з трьох пологів і близько 100 видів
- Гінкго: єдиний вид, який виживає лише там, де культивується
На додаток до цих п'яти збережених груп, існує кілька груп насіннєвих рослин, добре представлених у скам'янілостях, але більше не присутні. Ці вимерлі види іноді згортаються як «насіннєві папороті», але і грудості, і позначення як групи не вважаються точними: вони не тісно пов'язані з папоротями і, ймовірно, являють собою поліфілетичну угруповання. Більшість працівників вважають, що насіння еволюціонували неодноразово, і тому не повинно бути філогенетичної сутності, що відповідає ні насіннєвим рослинам, ні голонасінним (тобто насіннєвим рослинам, яким не вистачає квітів), хоча ці категорії зберігаються. Ми розглянемо подробиці життєвого циклу, зокрема форму жіночого гаметофіта і механізми, пов'язані з запиленням і заплідненням, для п'яти груп збережених насіннєвих рослин.
ТЕМИ
- Розвиток насіння хвойних порід
- Розвиток насіння в інших голонасінних
- Розвиток насіння у покритонасінних рослин
- Квіти
- квіткові модифікації
- Фрукти
Хвойні

Сосни - це найбільш часто зустрічаються хвойні породи та група, яка буде описана нижче, але основна модель тримається для всієї групи. Рослини, які визнані соснами, - це диплоїдні, спорофітні рослини. Всі хвойні дерева, включаючи сосни, гетероспористі і виробляють два види суперечок, обидва на одному дереві. Місцями виробництва спор є шишки. Шишки, які більшість людей визнають «сосновими шишками», - це жіночі соснові шишки. Це не тільки місце виробництва мегаспор, але і місце проростання мегаспор з утворенням гаметофіту, виробництва яєць цим гаметофітом, запліднення яєць та розвитку насіння. Всі ці події відбуваються в місці, описаному як яйцеклітина. Ці процеси, як правило, займають кілька років, і структури, які зазвичай визнають сосновими шишками, живуть і розвиваються протягом двох років або більше, причому багато значущих подій відбуваються, коли шишка набагато менша і не так легко спостерігати (рис. 1). Жіночий конус складається з осі (стебла), що несуть лусочки, піднесені приквітками, при цьому лусочки вважаються похідними змінених гілок. На верхній поверхні луски знаходяться яйцеклітини, структури, які переростають в насіння.

Чоловічі спори виробляються в менш звичних, але, безумовно, легко спостерігаються, чоловічих соснових шишках, які ростуть швидше, ніж жіночі шишки, але присутні на дереві набагато коротший час. Як правило, вони виробляються восени і видно у вигляді скупчення структур біля основи бутона. Вони розширюються навесні/на початку літа (рис. 2) і висихають і в'януть через місяць. На відміну від жіночих шишок, m ale шишки прості за будовою: гілка з щільно упакованими споровими несучими листям (спорофілами), кожна з яких має пару відносно великих спорангій на нижній поверхні.
Покоління гамет офітів розвивається з мікроспор і мегаспор, що виробляються і утримуються в чоловічих і жіночих шишках. Гаметофіти сильно скорочені і, особливо для самки гаметофіти, багато в чому невидимі через своїх невеликих розмірів і розташування. Чоловічі гаметофіти виробляються в мікроспорангії чоловічих шишок. Вони спочатку містять клітини, які піддаються мейозу для вироблення мікроспор. Спочатку спори знаходяться в скупченнях по чотири, що відображають своє походження в двох відділах мейозу (від однієї клітини до двох клітин до чотирьох клітин). Спори з часом відокремлюються і проходять дуже обмежений період розвитку, виробляючи гаплоїдний організм з чотирма ядрами, як правило, в трьох клітині (тобто одна клітина має два ядра), і володіючи двома крилоподібними повітряними мішками (рис. 3). Особливо значущим для розвитку пилкового зерна (він же чоловічий гаметофіт), є той факт, що його розвиток затримується. Це, і те, що мікроспорангіум розкривається, дозволяє пилку розсіюватися вітром.
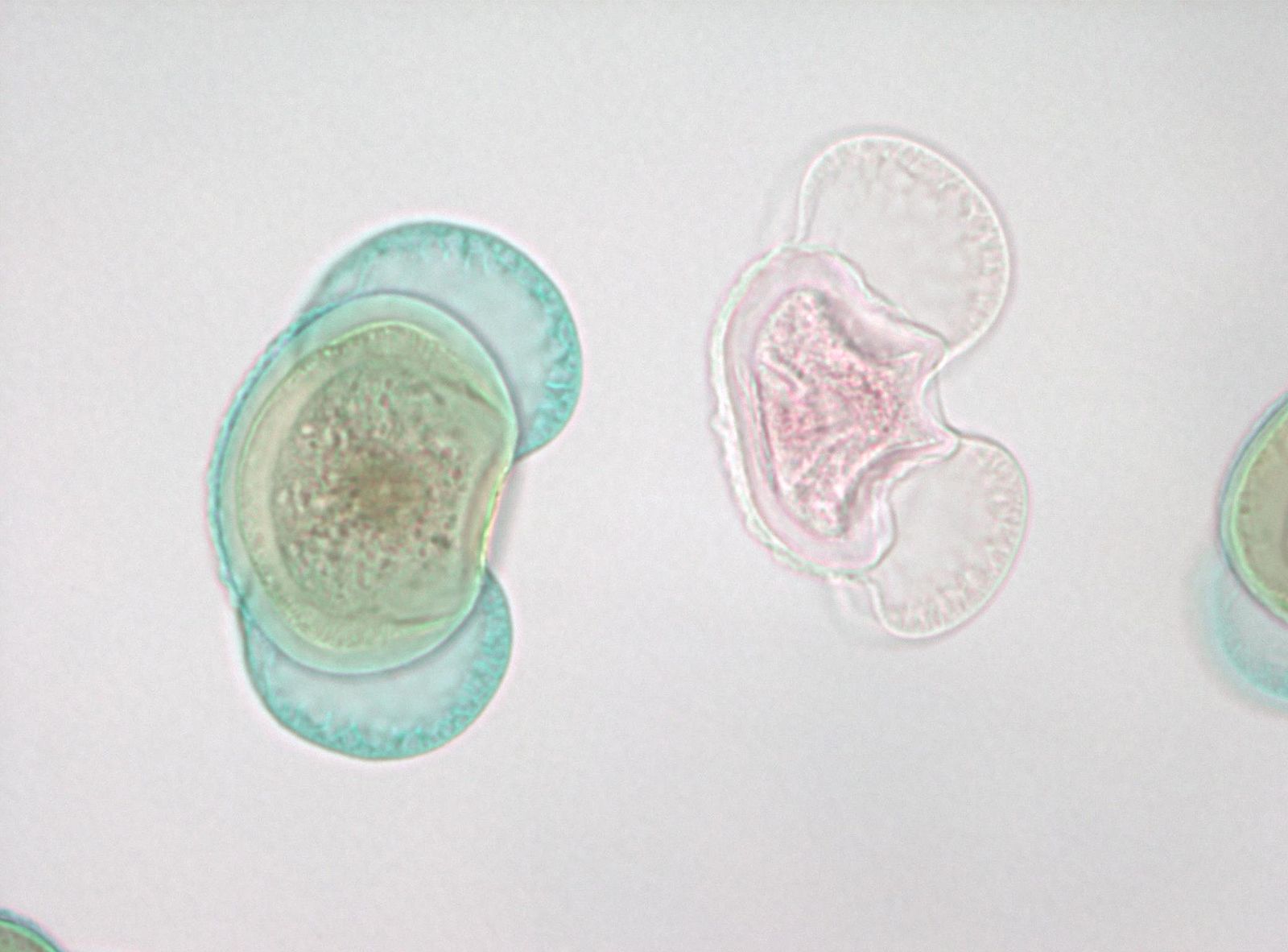
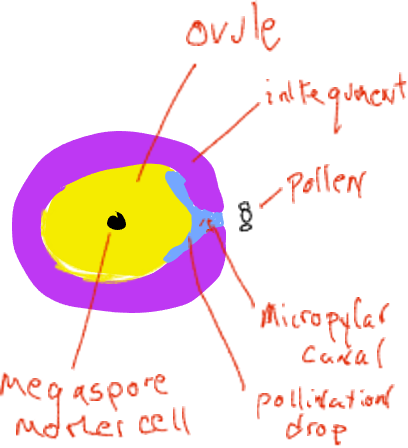
Запилення - це назва перенесення чоловічого гаметофіта (пилку), звідки він виробляється до місця розташування жіночого гаметофіта, у хвойних - рух від чоловічої соснової шишки до місця всередині жіночої шишки. У момент виділення пилку жіночі шишки дуже малі і «відкриті» з прогалинами над кожною окремою шкалою конуса, які відкриті назовні (рис. 4). Кожна жіноча конусна луска несе на своїй верхній поверхні дві яйцеклітини, кожна з яких має мегаспорангіум, вбудований в спорофітну тканину, звану покривами. На початку розвитку яйцеклітини є отвір, мікропілярний канал, між покривами, який з'єднується з простором між лусочками конуса. Пилкові зерна (чоловічі гаметофіти) в повітрі можуть ковзати між жіночими конусними лусочками і відкладатися в просторі поруч з мікропілярним каналом. Яйцеклітина виділяє рідку «краплю запилення» у цей простір, а пилкові зерна потрапляють у рідину та регідратують. У механізмі, не повністю вивченому, рідина з пилковими зернами виводиться через мікропілярний канал в простір на внутрішній стороні покривів, прилеглих до мегаспорангія.

Незабаром після цього покриви ростуть, щоб перекрити мікропілярний канал, і лусочки конуса ростуть, щоб закрити жіночий конус зовні. У цей час мегаспорангіум має єдину материнську клітину мегаспор, призначену піддаватися мейозу. Після мейозу лише одна з чотирьох дочірніх клітин залишається мегаспорою. The e megaspore не розпорошується, а розвивається в межах мегаспорангії в жіночий гаметофіт з декількох тисяч клітин, який, як правило, виробляє дві-три архегонії, кожна з яких виробляє одну яйцеклітину. Все це відбувається всередині жіночого конуса, який прикріплений і входить до складу спорофітної рослини. Жіночі гаметофіти хвойних дерев - це сильно знижені «організми», що знаходяться в овуляційних шишках, вбудованих всередину спорофітної тканини рослини.
У момент запилення зазвичай немає самки гаметофіта, тільки мегаспора. Усередині конуса самці гаметофіти продовжують свій розвиток, нехай і дуже повільно у сосн, вимагаючи 12 місяців між запиленням і заплідненням.
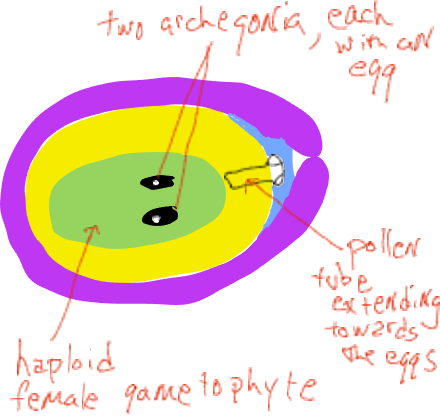
Оскільки мегаспора виробляє спору, яка повільно переростає у жіночий гаметофіт, вона проходить «вільну ядерну» стадію, де мітотичні відділи не супроводжуються утворенням клітинної стінки (організм коеноцитарний). Зрештою клітинні стінки утворюються, а жіночий гаметофіт утворює структури, описані як архегонія, кожна з яких має одну яйцеклітину.
У міру розвитку жіночого гаметофіту пилок проростає і одна клітина подовжується від зерна, зростаючи через мегаспорангіум (нуцелус) у напрямку до жіночого гаметофіту (рис. 6). Трохи більше року після запилення його трубчаста клітина зливається з яйцеклітиною і виділяються два ядра сперми, одне з яких зливається з ядром яйцеклітини, утворюючи зиготу, а інше ядро розпадається. За час між запиленням і заплідненням жіночі шишки виростають лише в невеликій кількості і вони залишаються закритими назовні. Зверніть увагу, що плаваюча сперма не виробляється, чоловічий гаметофіт виростає до яйцеклітини за допомогою подовженої клітини.
Після запліднення зигота розвивається в ембріон, вбудований і живиться жіночим гаметофітом. Тканини, що оточують жіночий гаметофіт, розвиваються в насіннєву оболонку, часто утворюючи структуру крила, яка дозволяє насінню розсіюватися вітром. У міру розвитку насіння шишка, що оточує його, також розвивається, часто істотно зростає. Після запліднення насіння може дозріти за короткий час, як і один рік, але для більшості видів це два роки або довше. У більшості сосен шишки з часом знову розкриваються, дозволяючи насінню випасти і розійтися вітром. Іноді шишки залишалися закритими і відкритими лише після сильної спеки багаття. Жіночі шишки ялівцю та тису розвивають плодоподібні риси, які приваблюють тварин, які полегшують розгін насіння, споживаючи «плоди» та випорожнюючи насіння на новому місці.
Інші голонасінні
Три інші групи насіннєвих рослин без квітів, Гнетофіти, Цикади та Гінкго демонструють ту саму основну схему виробництва насіння: чоловічі спори розвиваються в пилкові зерна, які диспергуються від спорофіту, щоб закінчити свій розвиток у структурі, яка виробляє жіночі спори, а отже, і жіноча. гаметофіти. Запилення принаймні у деяких цикад і у деяких гнітофітів включає комах; у гінкго і більшості гнітофітів запилення відбувається вітром. Gymnoserm буквально означає «голе насіння», і одна особливість, яка об'єднує неквітучі насіннєві рослини, полягає в тому, що під час запилення яйцеклітини доступні, а не поховані в тканині, через які повинен проростати чоловічий гаметофіт; натомість яйцеклітини доступні, принаймні на короткий проміжок часу, тому що конусні лусочки не зрослися один з одним і мікропілярний канал відкритий. Однак чоловічий гаметофіт, як правило, повинен рости через мегаспорангіум (нуцелус), щоб досягти яйцеклітини. У цикад самець гаметофіт насправді розвиває тип структури харчування (званий гаусторіум), розгалужену нитчасту структуру, що пронизує ядро і, мабуть, отримує від нього харчування. Врешті-решт джгутикова, рухлива сперма вивільняється і плаває через рідину «архегоніальної камери», області рідини між нуцелюлами та жіночим гаметофітом. Джгутикові сперматозоїди також містяться в гінкго.
квітучі рослини
Основний процес розвитку насіння у квітучих рослин такий же, як і у хвойних. До основних відмінностей можна віднести наступне:
- Чоловічі (виробляють пилок) і жіночі (виробляють насіння) органи, як правило, знаходяться разом в одній структурі, квітці, не розділені на двох різних гілках, оскільки вони знаходяться в хвойних порід.
- Яйцеклітини виробляються всередині структури, яка називається яєчником, яка не відкрита назовні, тим самим вимагають, щоб чоловічий гаметофіт рости через значну відстань спорофітної тканини, щоб контактувати з жіночим гаметофітом.
- Перенесення пилку (запилення) часто включає біологічні агенти (комахи, птахи, рідше ссавці) і різноманітні квіткові особливості підсилюють запилення.
- Жіночий гаметофіт, який називається ембріонним мішком (рис. 7).

Малюнок 7 Покритонасінна яйцеклітина, що містить зрілу самку гаметофіту (він же ембріонний мішок). Гаметофіт містить лише сім клітин, одну яйцеклітину, яка запліднюється, утворюючи зиготу, і велику центральну клітину, яка запліднюється, утворюючи триплоїдну ендоспермову тканину. Навколишні покриви розвиваються в насіннєву оболонку. оскільки вона з часом містить ембріон, ще більшескорочується, ніж у інших насіннєвих рослин, кардинально так, як правило, складається всього з семи клітин, шести гаплоїдних клітин, одна з яких - яйцеклітина, і одна більша центральна клітина з двома гаплоїдними ядрами.
- Обидва сперматозоїди, що виробляються чоловічим
гаметофітом, беруть участь у події запліднення (сингамії). Один зливається з яйцем, утворюючи зиготу, а другий зливається з центральною осередком. Це ядро сперми поєднується з двома ядрами центральної клітини, утворюючи триплоїдне ядро ендосперму. Потім центральна клітина розмножується, утворюючи тканину, ендосперм, який має обмежений розвиток і виявляється лише під час розвитку насіння і часто, але не завжди, у зрілому насінні. Початкові етапи розвитку ендосперму включають «вільно-ядерну» стадію, коли ядра діляться без утворення клітинної стінки, створюючи багатоядерну, всю триплоїдну (коеноцитарну) клітину. Цей матеріал називається «рідким ендоспермом» і знайомий як кокосове молоко, яке насправді є цитозолем. - У покритонасінних рослин жіночий гаметофіт, який дуже обмежений як за розміром, так і за тривалістю життя, не є живильною тканиною для ембріона, що розвивається, як у інших насіннєвих рослин, наприклад, хвойних. У покритонасінних рослин живильною тканиною для розвивається ембріона є ендосперм, тканина, що утворилася в результаті другої події запліднення.
квітка

Квітка являє собою сильно модифікований стебло, як правило, з чотирма мутовками, які згуртовані близько один до одного на кінці гілки. Вважається, що компоненти кожної з чотирьох мутовки представляють модифіковані листя, причому внутрішні дві мутовки - це сильно модифіковані спороносні листя (спорофіли) (рис. 8-9).. Елементи крайньої мутовки (чашолистки) самі листоподібні, хоча часто досить дрібні. Елементи наступної мутовки (пелюстки) часто мають листоподібну форму, але зазвичай представляють собою великі і барвисті структури, яким не вистачає хлорофілу. Наступна мутовка - це тамени, які часто складаються з стебла (нитки), що закінчується структурою, яка називається пильовиком , в якому виробляється пилок. Спочатку пильовики володіють мікроспорангіями, що містять мікроспорові материнські клітини. Вони виробляють мікроспори шляхом мейозу, і ці спори проростають і розвиваються у чоловічих гаметофітів, які називаються пилковими зернами, що складаються лише з двох або трьох клітин. Коли пилок дозріває, пильовик зазвичай відкривається, щоб зробити пилок доступним для запилювачів або вітру. P істили є найвнутрішнішою мутовкою і часто зрощені разом, тому центральна структура квітки - це єдиний (складний) маточок. Маточки (и), як правило, складаються з збільшеної основи (зав'язі), з його верхівки виходить стеблоподібна структура (стиль), яка закінчується поверхнею (рильцем), яка отримує пилок. Усередині зав'язі виробляється занадто багато яйцеклітин, які з часом стають насінням. До цього яйцеклітини є ділянками виробництва мегаспор, розвитку жіночого гаметофіту (= ембріонний мішок), запліднення і, нарешті, процесу розвитку насіння. Після запилення двох-триклітинний чоловічий гаметофіт (пилок) проростає на рильце і росте через стиль, а потім отримує доступ до яйцеклітини, зростаючи через мікропілярний канал, отвір між покривами які оточують кожну яйцеклітину. Зростання пилкової трубки передбачає розширення однієї клітини, яка називається пилковою трубкою, яка доставляє дві чоловічі гамети до кінця ембріонного мішка з яйцеклітиною . Трубчаста клітина зливається з ембріонним мішком і доставляє два сперми, щоб забезпечити подвійне запліднення, дозволяючи для подальшого розвитку зиготи та ендосперму.

квіткові модифікації
Варіації основного плану квітів - одна з дивовижних історій ботаніки і всієї біології. В цілому зміни можна віднести до сил природного відбору, що діють на взаємодію між запилювачами і рослинами. Деякі з поширених перетворень з описаного вище шаблону включають:
- злиття частин мутовки, наприклад, всі пелюстки зрощені разом, утворюючи чашку або воронку
- злиття членів двох мутовків (наприклад, злиття тичинок на пелюстках)
- зменшення кількості членів мутовки, зокрема скорочення від багатьох маточок до одного маточки, як це зустрічається у квіток квіток айстролисткових
- переходити від радіальної симетрії (всі частини мутовки мають однаковий розмір і орієнтовані подібним чином, рис. 9-10) до двосторонньої симетрії з квітами, що мають дві сторони, які є дзеркальним відображенням один одного (рис. 11), або іноді взагалі не мати симетрії.
- розміщення яєчника нижче місця прикріплення інших мутовки
- усунення множинних частин, іноді утворюючи одностатеві квітки (рис. 12).



Фрукти
Після запліднення яйцеклітина перетворюється в насінняв процесі, який передбачає узгоджений розвиток трьох різних тканин:
- зигота переростає в ембріон;
- ендосперм розростається спочатку за «вільно-ядерним» малюнком (ядерні поділи не супроводжуються утворенням клітинної стінки), а згодом, виробляючи нові клітини, і, нарешті, в деяких групах ендосперм зникає, коли ембріон збільшує свої сім'ядолі;
- тканини, що оточують ембріон і ендосперм, розвиваються в насіннєву оболонку.
Зауважте ще раз, що генетичний склад цих трьох компонентів відрізняється - ембріон є «новим поколінням» і є диплоїдним, ендосперм триплоїдний, а навколишні тканини диплоїдні, але є поколінням перед ембріоном.
Поки яйцеклітина перетворюється на насіння, зав'язь, а іноді й інші тканини, що оточують яєчник, розвиваються в структуру, яка називається плодом. Перетворення зав'язі в плід, як правило, передбачає вироблення нових клітин, зростання цих клітин і розвиток особливостей, характерних для спостерігаються рослини. Плід, як правило, має особливості, що посилюють розсіювання насіння і часто має функції, що захищають насіння. Хоча, як правило, легко відрізнити насіння від плодів, іноді між ними немає явної позначки, або може бути демаркація, яка обманює. Наприклад, мигдаль отримують з плоду, схожого на вишню (насправді вишня і мигдаль дуже тісно пов'язані). Їмо м'ясисту частину вишні, що входить до складу плодів. «Яма» насправді не є насінням, а скоріше це насіння, оточене найпотаємнішим шаром плоду. Коли ви «оболонки» мигдалю, ви розтріскуєте відкриту тканину плодів, щоб виявити одне насіння всередині. В цьому випадку захисну роль насіннєвої оболонки взяла на себе частина плодів (рис. 13). Аналогічно, соняшникове «насіння» насправді є однонасіннєвим плодом, і обстріл «насіння» фактично розщеплює плоди (рис. 14).