1.16: Репродукція - розвиток та фізіологія
- Page ID
- 7110

Репродуктивна/сексуальна поведінка і життєдіяльність організму

Навіть для найпростіших одноклітинних організмів розмноження вимагає зсуву «нормальної» діяльності. «Клітинний цикл» (рис. 1) описує закономірність діяльності, через яку клітини проходять з метою розмноження (тобто піддаються мітозу). У міру проходження клітиною цього циклу її діяльність змінюється. Зміни експресії генів призводять до змін від діяльності «росту та синтезу» (стадії G1, S, G2) до мітозної діяльності. Ще більше змін потрібно мати місце, якщо одноклітинний організм повинен брати участь в сексі. У якийсь момент статевого циклу мейоз повинен бути індукований, вимагаючи від клітин різко реорганізувати себе таким чином, що вдвічі зменшить кількість хромосом дуже специфічним чином. У деяких інших точках необхідно виробляти клітини з унікальними здібностями, здатністю зливатися один з одним.
Для багатоклітинних організмів необхідні зміни для розмноження або спричинення статевого процесу зазвичай включають глибокі зміни. Нові структури можуть з'явитися і поведінка багатоклітинного організмучасто зміщується, іноді різким чином, способами, які можуть навіть закінчити життя цього конкретного організму. Шаблони та контроль цих розробок розглянуті в цьому розділі.
ТЕМИ
- Відтворити живі структури: моделі виробництва в часі і просторі
- Контроль розвитку статі та розмноження - загальні сигнали
- Схеми розмноження у квітучих рослин
- Фотоперіодизм

Відтворити живі структури: моделі виробництва в часі і просторі
Для більшості організмів нормальна діяльність передбачає придбання речовини та енергії, а акт відтворення зміщує ресурси від структур, які набувають матерію та енергію, до тих, які беруть участь у відтворенні. Для плазмодійної слизової форми зрушення структури та поведінки є драматичним і охоплює весь організм: коеноцитарна плівка, яка поширюється по субстрату, знаходячи їжу, раптом переходить на зовсім іншу структуру, таку, яка нерухома і не в змозі харчуватися, але здатна виробляти пропагує.
Для інших організмів процеси як статі, так і розмноження передбачаютьконкретні структури, які безперервно або періодично виробляються, які спеціалізуються для цих цілей. Наприклад, нормальний ріст грибів виробляє гіфи, які здатні досліджувати середовище, в якому він росте, і отримувати матеріали, необхідні для росту. Щоб розмножуватися безстатевим шляхом, наприклад, з конідіями, грибкова нитка росте в іншому напрямку (зазвичай вгору) і виробляє ряд дрібних, легко відокремлюваних клітин, які можуть бути розпорошені. Таким чином, гриб змістив частину своєї діяльності з нормального режиму росту на репродуктивний режим. Іншим прикладом можуть бути диморфні листя у папоротей (рис. 2, див. Також чутливий папороть) та хвощів; рослина робить один вид листа для розмноження та інший тип листя для фотосинтезу - отже, його здатність до фотосинтезу знижується у міру збільшення його здатності до розмноження.
Для деяких рослин статеве розмноження передбачає драматичну перебудову, порівнянну зі слизовою цвіллю. Пам'ятайте, що квітка - це перетворена гілка і що гілки - це «нові модулі», важливі не тільки для їх росту, але й через потенціал виробляти ще більше модулів. Перехід цієї гілки до квітки змінює все це - вона перетворює індетермінантну структуру, здатну вирощувати та виробляти більше модулів, у детермінантну структуру, яка виробляє квітку чи квіти, а потім плід, а потім гине, усуваючи цей модуль та видаляючи його потенціал для виробництва будь-якого іншого модулі. Якщо всі пагони рослини переходять в цвітіння одночасно, торослина в цілому стає детермінантним і загине після плодоношення. Ось що відбувається в пшениці (рис. 3).

Ми можемо виділити дві «крайності» у схемі розподілу ресурсів для відтворення: одна передбачає поділ організму у просторі, а інша - розділення часу. Деякі організми, як папороті кориці, більшість водоростей і багато грибів, одночасно виробляють частини для розмноження і частини, які ведуть «нормальну» діяльність. Хоча ступінь розбиття може змінюватися з часом, організм росте і розмножується більш-менш безперервно. З іншого боку - організми, які проводять частину свого життя, ростуть, а потім перетворюються на інший тип організму, щоб розмножуватися. Приклади включають метеликів, метеликів, моркви (мережива королеви Анни), лопуха, буряка та коров'яку, всі з яких дають дуже короткий стебло з множинними листям і без гілок в перший рік життя. На другий рік стебло витягується, утворюються гілки і утворюються квітки і плоди. Велика частина матеріалу, що зберігається з першого року зростання, направляється на розвиваються насіння. Крім того, листя демонтують, щоб забезпечити додаткові ресурси. Отже, здатність рослини до фотосинтезу зникає і рослина гине в процесі цвітіння і плодоношення. У перший рік рослина орієнтується на зростання (придбання матеріалів), у другий - на розмноження.
Контроль розмноження і статі
У більшості організмів здатність до розмноження контролюється сигналами з навколишнього середовища, які відчуваються і спричиняють зміни в структурі та розподілі ресурсів. Хоча часто передбачається, що відтворення (а отже, і зростання чисельності населення) продиктовано ресурсами, безумовно, є ситуації, коли ресурси доступні, але організми не використовують ці ресурси для відтворення. Замість того, щоб бути прив'язаним до ресурсів, розмноження багатьох організмів пов'язане з конкретними сигналами в навколишньому середовищі - вони реагують на ці сигнали, відтворюючи. Зазначено інший спосіб, хоча для відтворення можуть знадобитися адекватні ресурси, їх часто недостатньо для відтворення. Крім усього іншого, це ускладнює прогнозування зростання населення. Ця тема буде розглянута докладніше при обговоренні поведінки популяцій. Тут ми обговоримо конкретні сигнали, які організми використовують для запуску репродуктивного процесу, сексуального чи безстатевого, а потім розглянемо (насамперед статеві) репродуктивні моделі, виявлені у квітучих рослин, та деякі специфічні сигнали, які контролюють репродуктивні моделі багатьох рослин.
Підказки для ініціювання розмноження та/або сексу
Зростання/розвиток т
Для багатьох одноклітинних організмів, що розмножуються мітозом, розмноження просто передбачає досягнення певного етапу клітинного розвитку. Це взагалі якось пов'язане з ростом, тобто накопиченням достатньої кількості матеріалів, щоб клітина могла бути розділена надвоє. Зростання пов'язаний з часом («клітини повинні досягти певного віку для розмноження»), але лише як опосередкований матеріальним придбанням. Бактерії, які діляться кожні 20 хвилин, не визначають проміжки часу, є процеси, що відбуваються, деякі з них принципово пов'язані з придбанням матеріалів, які займають 20 хвилин, щоб відбутися. При зміні температури або зміні доступності матеріалів бактерій буде потрібно більше часу, щоб розділити. Також, як зазначалося вище, організми мають здатність змінювати свою швидкість матеріального придбання, тобто вони контролюють, наскільки швидко вони ростуть, і через це вони можуть контролювати свою швидкість розмноження.
Час
Хоча всі еукаріоти, здається, вміють тримати час, дуже мало (? жодного?) мати відтворення або стать cued конкретно за часом (тобто мати секундомір, який ініціюється, а потім відтворюється в кінці певного часу). Однак звичайно, особливо для рослин, обраних для вирощування, що інтеграція часу та температури визначає цвітіння. Рослинам потрібно вирости до певної стадії, щоб розмножуватися, і для цього потрібен час і сприятливі температури. Багато пакетів насіння для домашніх садівників вказують щось на кшталт «квіти за 60 днів» - це припускає «нормальний» температурний режим; якщо температура холодна, це може зайняти 70 днів. Ця ідея інтеграції часу та температури розглядається докладніше в главі 26.
поживні речовини
Як обговорювалося вище, харчування (тобто придбання ресурсів, необхідних для зростання) явно може бути фактором, що впливає на ріст і розвиток, але іноді харчування відіграє більш конкретну роль. Хламідомонада секс викликається низьким рівнем азоту; розмноження fung al часто вимагає специфічної формулювання середовища, відмінного від середовища, що призводить до зростання; слиз цвілі можуть бути індуковані до споруляції (фаза статевого розмноження) конкретними режимами поживних речовин. Здатність деяких багаторічних рослин реагувати на квітучі сигнали була пов'язана з співвідношенням вуглецю/азоту в рослині, на які можуть впливати як фотосинтетична активність, так і наявність поживних речовин.
Несприятливі/вигідні умови
Дивно, але як хороші, так і погані умови можуть спровокувати стать або розмноження, залежно від виду. Ряд організмів змінюють свою діяльність, коли умови стають несприятливими (наприклад, високі або низькі температури, посуха, високий або низький рН, токсичні рівні певних хімічних речовин), і часто зміна передбачає розмноження та/або стать. Наприклад, плазмодії слизової цвілі (багатоядерні гігантські клітини) індуковані до формування спорангії, коли умови погані. Відомо, що ряд видів дерев цвітуть особливо добре, коли вони ось-ось загинуть. Таку поведінку можна вважати адаптивною, оскільки вона виробляє структури (насіння, спори), стійкі до суворих умов в той час, коли умови погіршуються. Такі споруди можуть бути вигідні ще й тим, що вони також передбачають розгін і пересування до нових, а можливо, і більш сприятливих, умов. У той же час припинення або скорочення виробництва пропаганди при несприятливих умовах може бути виправдано, оскільки ресурси необхідні для підтримки організму живим і не використовуються на «надмірності» розмноження та статі.
Підказки, які корисні для прогнозування майбутніх умов
Багато видів живуть у середовищах, які змінюються сезонно і мають певні частини року, які набагато сприятливіші для діяльності, пов'язаної з розмноженням/сексом, ніж інші періоди. Найбільш очевидною діяльністю, пов'язаною з розмноженням/сексом, є придбання матеріалу, але іншими факторами можуть бути: наявність агентів розгону (наприклад, вітру або, можливо, специфічних запилювачів або пожирачів фруктів), відсутність хижаків для потомства тощо Найбільш поширеним сигналом, який прогнозує майбутні умови, є фотоперіод. Фотоперіодичний контроль цвітіннябуде розглядатися після опису закономірностей репродуктивних зусиль, виявлених у квітучих рослин.

Схеми розмноження і статі у квітучих рослин
Покритонасінні рослини можна розділити на дві групи залежно від їх цвітіння: монокарпічні рослини, які цвітуть один раз і гинуть, і полікарпічні рослини, які цвітуть кілька разів. Монокарпічні рослини загинуть після цвітіння, оскільки всі їх меристеми були перетворені з індетермінантних вегетативних пагонів у детермінантні квітучі пагони. Після цвітіння квіти переростають у плоди, і рослина гине як тому, що більше немає вегетативних пагонів, щоб виробляти більше листя, і тому, що листя, які вже були присутні, були «відбиті» як джерело матеріалів для насіння, що розвиваються, і, отже, були знищені. Про це йшлося вище і зустрічається в ряді видів сільськогосподарських культур, включаючи кукурудзу, пшеницю, сою (рис. 3 і 4)
Багато монокарпічні рослини - однорічні, що живуть всього рік. Що б не знадобилося, щоб рослина цвіла, відбувається протягом року з моменту проростання насіння. Якщо задіяний конкретний кий, то протягом року рослина розвиває чутливість до кия і може відреагувати. Зауважимо, що деякі однорічні рослини цієї місцевості, особливо вирощені в садах, можуть бути однорічними тільки через усунення сприятливих умов восени - вони просто гинуть від морозу і холоду і жили б довше, якби умови були більш сприятливими. Більшість цих рослин не є рідними для цієї місцевості; більшість однорічних рослин, які є рідними для цієї місцевості, вбивають себе в процесі цвітіння. У помірних північноамериканських середовищах існування більшість однорічників проростають навесні і цвітуть влітку, але деякі, звані зимовими однорічними рослинами, проростають восени, зимують і цвітуть навесні. Більшість пшениці, вирощеної на півночі США, вважатиметься озимою однорічною, хоча в центральній та південні частини країни пшениця вирощується звичайна однорічна і висаджується навесні.
Монокарпічні рослини також можуть бути дворічними, іноді визначаються як рослини, які живуть два роки, але точніше описуються як рослини, які живуть протягом двох вегетаційних сезонів. Ці рослини, як правило, зустрічаються в місцях проживання, які є сезонними, тобто мають частину року, сприятливу для росту і частину несприятливу для росту, як правило, через низькі температури, але зрідка через брак вологи. Дворічні поводяться так, як вони роблять, оскільки протягом першого вегетаційного періоду вони не реагують на сигнали, що викликають цвітіння, тоді як у другому сезоні вони є. Прикладом може бути буряк s, вид, який потребує нас холодної зими, щоб мати можливість реагувати на сигнали (фотоперіод, див. нижче), які викликають цвітіння g Якщо він не цвіте перший вегетаційний період, хоча він отримує сигнал до квітки, але цвіте на другий рік після впливу холоду. Дворічні є монокарпічними і, як правило, демонструють значний диморфізм між першим і другим роком, часто мають дуже короткий стебло і без гілок перший рік, а подовжений і розгалужений стебло другий рік. Як згадувалося раніше, можна подумати про форму першого року, пов'язаного з придбанням ресурсів, а форма другого року пов'язана з відтворенням.
Хоча це рідкість, кілька монокарпічних рослин багаторічні, живуть кілька років, а потім цвітуть один раз (рис. 5). Приклади включають рослину століття (Agave), бамбук та деякі види тирлич. Випадок бамбука особливо важливий для ведмедів панд, оскільки вони харчуються майже виключно вегетативними (тобто неквітучими) рослинами бамбука, які можуть рости до 70 і більше років, утворюючи великий клон. Коли рослина цвіте, гектари бамбука, продукт 70+ років зростання, виробляє насіння (які панди не їдять) і в процесі цього бамбук гине, залишаючи панду без джерела їжі.

Полікарпічні рослини багаторічні. Вони потенційно живуть вічно, оскільки лише деякі модулі перетворюються на квітучі пагони в будь-який конкретний рік. Оскільки вся рослина піддається однаковому набору сигналів, різна поведінка різних меристем, деякі з яких виробляють квіти, а деякі ні, є результатом різної чутливості до сигналів. Поширеною схемою є те, що на квітучий кий реагують лише однорічні стебла (див. Мачу-й-мачуха). Таким чином, рослина матиме дві групи пагонів, пагони поточного року, які не реагують, і пагони попереднього року, які дійсно реагують, а тому дають квіти та плоди і гинуть. Значить, навесні року ці рослини складаються виключно з пагонів, які стануть квітами. Але перш ніж вони зацвітуть, вони виробляють гілкові пагони, які не стануть квітами до наступного року. Варіант такого малюнка спостерігається у багатьох сортів малини/ожини. Хоча рослина багаторічна, це результат підземних стебел. Вертикальні стебла, звані тростини, дворічні, перший рік ростуть вегетативно, другий рік дають квіти і плоди і гинуть. Малина пляма багаторічна, але стебла, які ви бачите, живуть лише два роки, кожна тростина поводиться як дворічна.
Квітучі рослини сигнали для розмноження
Хоча кілька рослин, особливо рослин, вибраних для вирощування, не потребують специфічного екологічного сигналу, щоб викликати цвітіння, вони просто цвітуть після того, як рослина досить виросла, більшість рослин вимагають конкретних екологічних сигналів, щоб викликати цвітіння. Однак для багатьох рослин одного кия недостатньо, щоб спровокувати цвітіння; сама рослина повинна вміти реагувати на кий, тобто рослина спочатку не реагує і не реагує на певні сигнали. З часом рослина розвиває чутливість до кисті і може реагувати, коли відбувається лікування. Таким чином, коли ми розглядаємо, що робить рослину квіткою, ми повинні враховувати можливість того, що задіяні два процеси: один запускає чуйність, а другий запускає саме цвітіння. Хорошим прикладом цього є цвітіння буряка: для того, щоб цвісти, вона повинна спочатку піддаватися впливу прохолодних температур протягом певного періоду часу; це розвиває його здатність реагувати на специфічний сигнал, який викликає цвітіння, тобто фотоперіод, особливе поєднання світла та дня в 24-годинний період. Буряк не реагує на фотоперіод, якщо вона вперше не піддалася впливу прохолодних температур. Отже, він росте перший рік без цвітіння, і лише протягом другого літа, після періоду прохолодних температур взимку, його спонукають до цвітіння.
Фотоперіодизм
Найрізноманітніші організми, включаючи рослини, тварини, гриби і протисти, реагують на фотоперіод, відносну кількість світла і темряви в 24-годинний період. Для багатьох організмів, включаючи багато рослин, він є одним з ключових детермінант, що впливають на розмноження (рис. 6). Спеціально для квітучих рослин фотоперіод часто визначає, коли квіти рослини і фотоперіод, ймовірно, найбільш широко вивчалися в цьому контексті. Але важливо розуміти, що фотоперіод часто впливає на фізіологію, структуру та поведінку і що його механізм дії знаходиться на молекулярному рівні, тобто впливаючи на те, які гени виражаються.
Фотоперіод є значним сигналом для організмів, що живуть на північ або південь від екватора, оскільки фотоперіод прогнозує майбутні умови. Наближається зима може бути відчута скороченням фотоперіодів; майбутню весну можна почути, подовжуючи фотоперіоди.
Рослини виявилися чудовими організмами для вивчення фотоперіодизму, оскільки для деяких з них один день певного фотоперіоду може призвести до вимірюваної реакції. На відміну від цього, для деяких організмів, та й для більшості рослин, реакція очевидна лише після тривалого впливу певних фотоперіодів.
Ранні дослідження встановили той факт, що саме нічний період є критичним. Переривання темного періоду може змінити реакцію, тоді як переривання світлового періоду не змінює поведінку. Здатність реагувати на фотоперіод вимагає двох здібностей: здатності відчувати світло проти темряви, тобто фотосенсор, і здатність тримати час. У всіх досліджуваних організмах механізм синхронізації, здається, пов'язаний з внутрішньою 24-годинною ритмічністю, щось описується як циркадний годинник, оскільки ритми в поведінці мають періодичність навколо (приблизно = навколо) 24 години. Циркадні ритми, ймовірно, зустрічаються у всіх еукаріотичних організмах, безумовно, вони, здається, є особливістю більшості еукаріотичних організмів, де його шукали.

Фактична фотоперіодична реакція є результатом певної картини світла і темряви, яка накладається на внутрішній 24-годинний ритм організму. Як результат, поведінка, що спостерігається після надання організму 12 годин світла та 12 годин темряви, залежить від того, яку частину його 24-годинного циклу застосовується 12 годин темряви. Механізм синхронізації - це не секундомір, який разів темний період, критично те, як періоди світла/темряви взаємодіють з внутрішніми ритмами, які мають 24-годинну періодичність. Зазначено інакше, існує періодичність реакції рослин на темряву.
Найбільш часто зустрічаються фотоперіодичні реакції включають розмноження, але загалом фотоперіод може бути організаційним фактором, що визначає закономірності росту та розвитку, які проявляє організм. Деякі з них перераховані нижче:
У квітучих рослин:
- квіткове виробництво
- температурна толерантність
- виробництво вегетативних бруньок

Малюнок 7 Картопля утворюється, коли кінчики коротких підземних стебел збільшуються і зберігають вуглеводи. Це перетворення являє собою фотоперіодичну реакцію, спровоковану більш короткими днями. - активізація бутонів
- активізація бічних бутонів
- формування бульб (рис. 7)
- пророщування насіння
У неквітучих насіннєвих рослин:
- виготовлення шишок
- формування і активізація бутонів
У динофлагелатах:
- утворення кісти
Як у червоних, так і в бурих водоростях:
- закономірність росту і формування репродуктивних структур
У тварин:
- розвиток яєчників і насінників у птахів та інших тварин і пов'язані з цим зміни в поведінці
Для квітучих рослин реакція фотоперіодичного цвітіння, як правило, поділяється на одну з трьох категорій:
- довгоденні рослини, які цвітуть тільки в тому випадку, якщо тривалість днів (періоди світла) довше деякого критичного значення.
- короткоденні рослини, які цвітуть тільки в тому випадку, якщо дні (періоди світла) коротше деякого критичного значення.
- денні нейтральні рослини, цвітіння яких явно не прив'язане до фотоперіоду
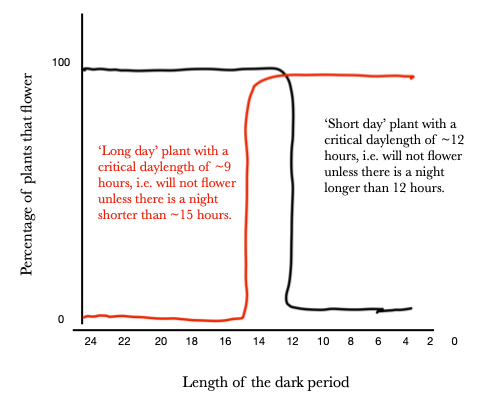
Див. Рисунок 8. Короткоденні рослини (чорна лінія) потребують нічних періодів довше, ніж якесь критичне значення, щоб цвісти. Довгоденні рослини (червона лінія) потребують довжини ночі коротше, ніж якесь критичне значення, щоб цвісти. Фактична критична тривалість дня (тобто вертикальна частина лінії) може зміщуватися вправо або вліво залежно від конкретного виду (або сорту всередині виду). Зауважте, що жодна з двох проілюстрованих тут рослин не буде цвітінням з тривалістю ночі між 15 і 12 годинами. Аналогічно, якби червона лінія була зрушена досить вправо, або чорна лінія досить вліво, виникли б ситуації, коли рослини як довгоденні, так і короткоденні цвітуть в один і той же фотоперіод.
Хоча назви стосуються періодів світла в 24-годинний період, насправді нічний період є критичним. Отже, рослини довгого дня краще назвати рослинами короткої ночі, а рослини з коротким днем краще назвати рослинами довгої ночі (але їх не так!!). Ще одним заплутаним фактором, який слід оцінити, є те, що хоча ми можемо вважати, що будь-який день з більш ніж 12 годин світла є «довгим днем» (і «короткою ніччю»), що є критичним є фактичне довжина в порівнянні з критичним значенням. Отже, можна цвісти як довгоденні, так і короткоденні рослини під одним фотоперіодом: фотоперіод 10 годин світла і 14 годин темряви спровокує цвітіння рослини довгого дня з критичним значенням 9 годин світла, а також короткоденна рослина з критичним значенням 11 годин світла.
Хоча реакції рослин потрапляють на ці три загальні типи, фактичні відповіді часто ускладнюються двома факторами. Одним з них є те, що реакція рослини на певне лікування може не обов'язково «все або нічого» (описана як «якісна реакція»), що означає, що рослина не цвіте, якщо не отримає відповідного стимулу. Для багатьох рослин реакція на відповідний стимул є «кількісною», що означає, що рослина цвіте швидше або з більшою кількістю квітів, якщо отримує певний стимул (лінії переходу на графіку вище можуть бути не вертикальними, а натомість мають нахил). Другим ускладнюючим фактором є те, що фотоперіодична чутливість може включати в себе кілька послідовних сигналів, включаючи ті, які не включають фотоперіод. Наприклад, деякі рослини реагуватимуть лише на довгі дні після коротких днів або реагують на довгі дні лише через певний проміжок часу в прохолодних температурних умовах.
У рослині з'являється два пігменти, фітохром і криптохром, які можуть взаємодіяти з циркадними ритмічними явищами і виробляти фотоперіодичні реакції. Хоча обидва ці пігменти взаємодіють з декількома різними фізіологічними процесами, реакція цвітіння, здається, є результатом змін експресії генів, що виникають внаслідок систем трансдукції сигналів, які впливають на взаємодію білка/ДНК. Зокрема, відповідний фотоперіодичний стимул, як видається, призводить до вироблення специфічних молекул мРНК, які важливі для перетворення верхівкових меристем вегетативних пагонів у квіткові меристеми.