1.13: Пол і розмноження у ненасіннєвих рослин
- Page ID
- 6953

Практично всі рослини здатні до розмноження без статі. Найчастіше це трапляється внаслідок того, що рослина розрізається на шматки, і ці шматки здатні регенерувати втрачені частини. Крім того, деякі види розробили шматки, які особливо схильні до відламування, наприклад, gemmae (однина від gemma) деяких мохів і печінки. Досить часто квітучі рослини виробляють безстатеві розмноження в місцях, де насіння нормально розвиваються, утворюючи насіннєподібні структури без нормального процесу сексу. Хоча розмноження без статі поширене у рослин, і для деяких видів воно є їх єдиним засобом розмноження, більшість рослин покладаються на статеве розмноження, і два нововведення, насіння та квітка, були особливо важливими для еволюції рослин.
Що стосується статі, рослинне царство ілюструє чудові закономірності єдності та диверсифікації. З одного боку, все рослинне життя демонструє принципово схожу закономірність статі, з іншого боку, рослини ілюструють неабияку диверсифікацію цієї загальної теми.
ТЕМИ
- чергування поколінь
- Мохи, печінка та рогач
- Папороті
- Хвощі
- Клубні мохи
Всі рослини проявляють чергування поколінь s, вони виробляють два типи багатоклітинних організмів: один диплоїдний і отриманий в результаті розвитку зиготи, і один гаплоїдний і отриманий в результаті розвитку гаплоїдної клітини, званої спорою. «Розвиток» передбачає проліферацію клітин, диференціювання клітин і формування специфічних структур з конкретними ролями, тобто морфогенез.
Всі рослини демонструють чергування поколінь і чергуються між гаплоїдними і диплоїдними організмами. Для завершення статевого циклу гаплоїдний організм повинен виробляти гамети, які об'єднуються, утворюючи зиготу. Тобто серед всіх гаплоїдних клітин, які виробляються в результаті клітинних поділів гаплоїдної спори, частина клітин наділена особливими можливостями, що дозволяють їм стикатися і взаємодіяти з іншою гаметою для утворення зиготи. Аналогічно, з усіх диплоїдних клітин, отриманих від зиготи, деякі, звані «споровими материнськими клітинами», наділені здатністю піддаватися мейозу виробляти гаплоїдні клітини, спори, які переростають у гаплоїдних особин.. Зверніть увагу, що мейоз НЕ виробляє гамети, скоріше він виробляє суперечки, які розсіюються і розвиваються в гаплоїдні рослини. Диплоїдна рослина, яка виробляє спорові материнські клітини та спори, називається «спорофітом». Організм, що виробляє гамет, який є гаплоїдним, називається гаметофітом. Таким чином, відбувається «чергування поколінь» з гаплоїдної формою, що чергується з диплоїдною формою (рис. 1). Часто одну з двох форм складно оцінити, багато в чому тому, що їх часто складно побачити - вони часто маленькі, недовговічні і можуть не жити окремим існуванням від попереднього покоління. Тобто гаплоїдні та диплоїдні форми можуть не просторово відрізнятися один від одного; вони можуть бути тимчасово, або постійно, частиною свого батька!
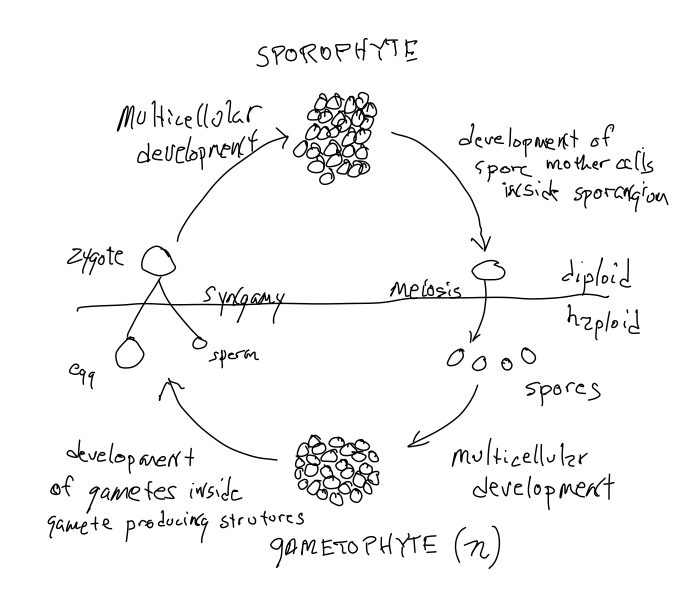
Як і у знайомих тварин, гамети всіх рослин бувають двох типів: більша, нерухома яйцеклітина та менша сперма, яка повинна певним чином бути рухомою, щоб зіткнутися з яйцеклітиною. Гаметофіти рослин можуть бути двостатевими (гермафродити), що продукують як яйцеклітину, так і сперму, або одностатевими, тобто існують окремі чоловічі і жіночі гаметофіти. У деяких групах рослин, включаючи всі ті, що обговорюються в цьому розділі, сперма рухлива, оскільки вони мають джгутики. Сперми відпливають від чоловічого гаметофіту, який його виробляв, і хімічно притягуються до структури, що виробляє яйцеклітини. Оскільки рослини є наземними організмами, вода, необхідна для того, щоб джгутиковий сперматозоїд підпливав до яйцеклітини, присутній не завжди. У наступному розділі ми побачимо, що деякі рослини обійшли цю проблему, змусивши чоловічих гаметофітів стати рухливими, а не спермою, яку вони виробляють.
Як і у тварин, самка гамети, яйце, не рухається. Вона зберігається в організмі, який її виробляє. У випадку з рослинами це гаплоїдний гаметофіт (не диплоїдна особина, як у ссавців). А у деяких рослин, розглянутих у цьому розділі, зигота, утворена заплідненням яйцеклітини, виростає з архегонії (структура, в якій виробляється яйцеклітина) і після запліднення виробляє спорофіт, прикріплений до гаметофіту. Як варіант, весь (жіночий) гаметофіт деяких груп рослин вбудований в диплоїдну рослину (спорофіт), яка виробляла спору, яка переросла в жіночий гаметофіт. Тобто в деяких групах рослин спори, що виробляються спорофітами, не розсіюються, натомість спора проростає там, де вона виробляється, і росте, утворюючи рослину гаметофіту на рослині спорофітів або всередині неї. Рухливі чоловічі гаметофіти та осілі жіночі гаметофіти - це особливості кількох рослин, розглянутих у цьому розділі, але набагато значно розвинені у насіннєвих рослин, розглянутих у наступному розділі. У цьому розділі ми розглянемо більш охоче спостережуване і зрозуміле чергування поколінь, яке зустрічається у рослин без насіння.
Мохи Печінкова і Роголистник

У мохів, печінкиі рогачів (трьох груп рослин, позбавлених судинної тканини) саме рослина гаметофіт живе найдовше і є найбільш помітним. Він виробляє форму, здатну збирати матеріали для росту, насамперед за допомогою фотосинтезу (рис. 2). Найпоширеніша форма, що зустрічається у всіх мохів і багатьох печінкових, - це подовжена вісь, що несе клапті тканини, що збільшують область фотосинтезу. У якийсь момент, часто раз на рік, гаметофіт виробляє гамети, що виробляють структури, які називаються архегонією (сингулярна = архегонія), якщо вони виробляють яйцеклітини та антеридії (сингулярні = антеридій), якщо вони виробляють сперму. Ці структури, як правило, виробляються посеред модифікованих «листя» на кінцях стебел/гілок у мохів або, у деяких печінкових вертушок, вони виникають на нижній стороні парасолькові/грибоподібних структур, які простягаються вгору від основного тіла талоїдної печінки.. Антеридії (рис. 3) - це приблизно сферичні контейнери, в яких клітини диференціюються на джгутикові сперми і виділяються, коли антеридії розриваються. Архегонія - це структури у формі колби з однією яйцеклітиною біля основи та подовженою шийкою, яка розвиває канал, через який сперма може плавати, щоб запліднити яйцеклітину. Отримана зигота розвивається всередині архегонії, виробляючи спорофіт, який в кінцевому підсумку простягається, як правило, на один-шість см за межі архегонії і існує як придатк гаметофіту, ніколи не живучи незалежним від нього існування. Хоча спорофіт іноді зелений і здатний до фотосинтезу протягом частини свого життя, його структура, простий стебло без «листя» для збільшення площі поверхні, не особливо підходить для фотосинтезу. Швидше структура підходить для розгону спор; стебло зазвичай піднімає капсулу, яка є місцем, де розвиваються спорові материнські клітини, піддаються мейозу та виробляють спори.


Якщо «просторове поділ» використовується суворо для визначення організму, то ці спорофіти не вважатимуться окремим організмом, вони просто є частиною гаметофіту (рис. 5). Але, в світлі статевого циклу і закономірності чергування поколінь, корисно вважати їх окремими організмами. Це підтверджується тим, що вони мають різну кількість хромосом, ніж гаметофіт, з якого вони виростають.

У всіх інших рослин, крім мохів, печінкових і рогоцвітів, саме рослина, що виробляє спор, живе найдовше і є найбільш помітною; це форма, яку ми бачимо і визнаємо як рослину. Т він спорофіт все ще починає свій розвиток, зростаючи з архегонії, де яйцеклітина вироблялася і була запліднена, але зростання спорофіта такий, що він стає повністю незалежним від гаметофіту, з якого вона виходить, і врешті-решт має повністю автономне існування. Таким чином, коли ми бачимо папороті, хвощі, клубні мохи та насіннєві рослини, ми спостерігаємо диплоїдну рослину, яка виробляє спори. У всіх цих групах гаметофіт невеликий і невловимий, але основний життєвий цикл такий же, як у всіх рослин: чергування поколінь між гаметофітом і спорофітом. Насіння і насіннєві рослини розглядаються в наступному розділі, нижче розглянуті аспекти статевого циклу, зокрема особливості рідше помічених гаметофітів, які виробляються безнасінними судинними рослинами.
Папороті
Більшість папоротей мають невеликий, фотосинтетичний гаметофіт, який зазвичай менше 1 см поперек і товщиною однієї клітини, т. Е. Він не має осі стебла і часто має «серцеподібну форму» (рис. 6-8 і 11). Зазвичай він прикріплюється до субстрату за допомогою ризоїдів (ниток нефотосинтетичних клітин).

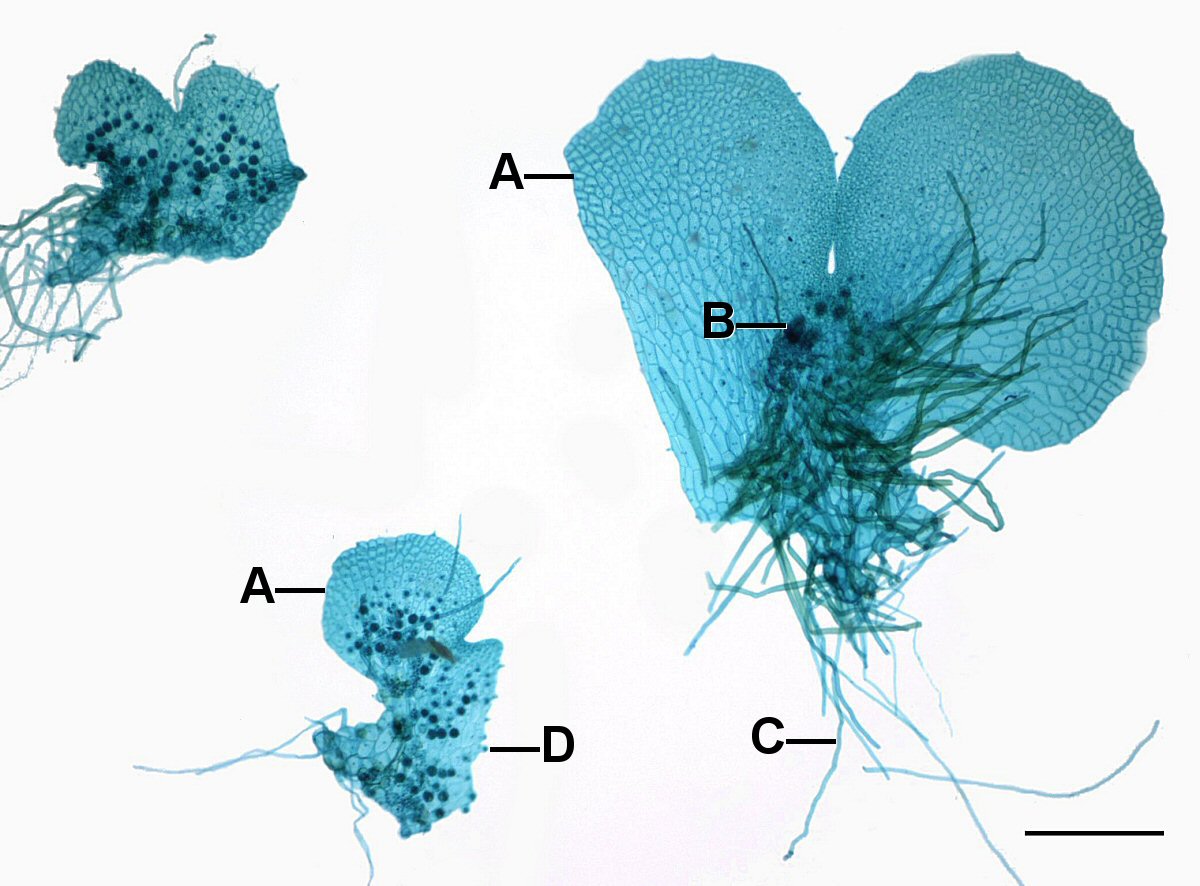
Як це було у мохів, гаметофіти папороті виробляють структури, де яйцеклітина та сперма виробляються в результаті ділення клітин за певною схемою для утворення архегонії (рис. 7-9) та антеридій. Важливо відзначити, що гамети не виробляються мейозом, оскільки всі клітини гаметофіту вже є гаплоїдними.



Гаметофіти папороті, як правило, мають колбоподібні жіночі структури (архегонії), розташовані в виїмці між частками і кулястими чоловічими структурами (антерідіями), розташованими на частках. Хоча більшість гаметофітів папороті є гермафродитними, деякі є одностатевими, а для деяких їх сексуальне вираження залежить від умов навколишнього середовища. Всі клітини гаметофіту є гаплоїдними, але він виробляє клітину, яйцеклітину, з особливими здібностями розвитку. Антеридії виділяють сперму, яка має джгутики, які дозволяють їм плавати до архегонії, де розташовані яйцеклітини, плавають по вузькому каналу і зливаються з яйцеклітиною біля основи. Зигота розвивається в спорофіт, виробляючи стебла і коріння. Стебла утворюють листя, які затінюють гаметофіт, і він незабаром відмирає (рис. 9)

Спорофіт продовжує рости, утворюючи папороть, який ми визнаємо. Він має таку ж будову, як і більшість рослин: корінь-вісь пагонів з листям, що виробляються втечею. Більшість папоротей в цій місцевості мають стебла (кореневища), які знаходяться під землею і відносно короткі. Те, що ми бачимо, - це листя, що виходять над землею з цьогокореневища. У якийсь момент цей диплоїдний організм виробляє структури під назвою спорангія, всередині яких знаходяться спорові материнські клітини, які піддаються мейозу, щоб виробляти групу з чотирьох (тетрада) гаплоїдних суперечок, які виділяються в навколишнє середовище. Коли вони проростають, вони переростають в гаплоїдні гаметофіти і процес повторюється. Як правило, спорангії виробляються в кластерах, які називаються «фруктовими крапками», які розташовані на нижній стороні листя (див. Деревний папороть). Інші папороті мають цілі частини листя, які очевидно відрізняються і де утворюються суперечки. Кілька папоротей у цій області є диморфними (див. Чутливий папороть), утворюючи два типи листя, деякі зелені та фотосинтетичні і які ніколи не виробляють спори та інші листя, які не є фотосинтетичними і виробляють рясні спори, при цьому харчуються фотосинтетичною частиною рослини.
Хоча це загальна картина для папоротей, існує певна варіація, одним із прикладів якої є водяний папороть Marsilea, який має кілька цікавих особливостей (див. Інформаційний лист про Marsilea). Як і у ряді папоротей, суперечки виробляються на спеціалізованому листі, який сильно відрізняється від звичайних фотосинтетичних листя. У той час як нормальні листя зелені і мають форму листя конюшини, листя, що несуть спори, спочатку упаковані в структуру, схожу на насіння, тверду зовні і здатні висушуватися та оживити (пророщувати) при повторному змочуванні. В цей час «плодоносний» (тобто спороносний) лист виходить у воду, дуже мало схожий на лист: він без хлорофілу, дуже дрібний і драглистий. Він виробляє спори в скупченнях, і існує два типи спор: чоловічі спори, які називаються мікроспорами, і жіночі спори, звані мегаспорами, кожен в окремій спорангії. Технічна назва рослин, які виробляють два види суперечок, - гетероспорие. На відміну від цього, більшість папоротей гомоспорні, утворюючи лише один тип спор, який зазвичай виробляє гермафродитні (бісексуальні) гаметофіти (описані вище); кілька гомоспорних форм виробляють одностатеві гаметофіти, як чоловічі, так і жіночі, але обидва походять від однакових на вигляд спор.
Два види спор Марсілеї легко розрізняються за розмірами. Мегаспори мають довжину близько 1 мм і проростають для отримання яєць, жіночих гаметофітів. Мікроспори мають довжину лише ~ 70 мкм і виробляють чоловічі гаметофіти, що виробляють сперму. Без будь-якого збільшення розміру і не виходячи з мікроспор, чоловічі спори проростають, утворюючи «рослину» з ~ 35 клітинами, 32 з них - сперматозоїди, багатобічеподібні сперматозоїди у формі штопора розміром приблизно 10 мкм, які вивільняються від чоловічого гаметофіту при порушенні стінки мікроспор. Сперматозоїди хімічно притягуються до жіночого гаметофіту. Жіночий гаметофіт істотно більше, ніж чоловічий гаметофіт, але він все ще невеликий і, як і чоловічий гаметофіт, проявляє ендоспоричний розвиток, його розвитоквідбувається в межах спорового випадку мегаспори, при цьому від нього відходить лише дуже коротка шия. Він виробляє єдиний архегоній з однією яйцеклітиною, до якої сперматозоїд плаває і запліднює, утворюючи зиготу. У той час як нова спорофітна рослина, здавалося б, проростає з жіночої суперечки, вона насправді походить від жіночого гаметофіту, який росте всередині спорового корпусу.
Подальше читання
«Marsilea: середовище проживання, зовнішні особливості та розмноження» на Біологія Discussion.com
Ще один цікавий папороть - папороть з щетиною Аппалачі, який відомий лише з гаметофітної форми. Мабуть, він розмножується безстатевим шляхом мільйони років! Існує кілька інших видів папоротей, відомих лише як гаметофіти.

Хвощі
Основний малюнок, виявлений у папоротей, з домінуючим поколінням спорофітів та зменшувальним поколінням гаметофітів, знаходиться в хвощах, групі судинних рослин, які виникли в палеозої і виробляли ряд деревних форм, які були значними для отримання великих родовищ, які стали вугіллям і масло. Залишається лише один рід хвощів, а у всьому світі налічується менше 20 видів. Всі вони трав'янисті з багаторічними кореневищами, які посилають вертикальні гілки, які мають дуже характерний малюнок росту з фотосинтетичними стеблами, дуже дрібними лускоподібними листям та мутовчастими гілками або без гілок. Спори виробляються в термінальній конусоподібної структурі, що представляє собою скупчення спорангії. Спори розсіюються вітром, але їх руху та виходу зі спорангії можуть сприяти структури, звані елаторами, ремінцеподібними придатками на спорах, які рухаються у відповідь на поглинання та втрату води. Проростання спори дає невеликий (~ 1-3 см) фотосинтетичний гаметофіт, схожий на штифтову подушку. Дуже короткі «стебла» присутні з придатками, які збільшують фотосинтетичну площу. Антеридії та архегонія зазвичай виробляються з одного гаметофіту, хоча він може бути одностатевим протягом певного періоду часу. Сперматозоїди мульти-джгутикуються і потрібно плавати, щоб дістатися до яйцеклітини. Запліднення призводить до зиготи, яка розвивається в диплоїдний спорофіт, який незабаром заростає гаметофіт, з якого він виходить, утворюючи коріння і як горизонтальні стебла (кореневища), так і вертикальні стебла.
Клубні мохи, колосисті мохи та перо
Вважається, що три групи є тісно пов'язаними і згруповані разом як «лікоподи» або «лікофіти», група, яка також включає ряд викопних форм, включаючи деревоподібні форми, які були дуже важливими наприкінці палеозойської ери. Група налічує 1200 видів і значно різноманітніша за хвощі, але набагато менш різноманітна, ніж папороті (12 000 видів). Спорофіти збережених клубнихмохів, колосових мохів та перо - все це трав'янисті багаторічники. Зазвичай вони широко поширюються надземними та підземними стеблами (тропічні члени, як правило, епіфіти). Хоча стародавні члени групи демонстрували деревне зростання, жоден з видів, що живуть сьогодні, не роблять.
Клубні мохи (рис. 13) унікальні тим, що мають гаметофіти, які є підземними та нефотосинтетичними, виживаючи як паразити, харчуючись грибами. Ці гаметофіти живуть набагато довше, ніж більшість гаметофітів судинних рослин, деякі більше 15 років. Колосові мохи та перо гетероспористі, і, як і водний папороть Marsilea, гаметофіти розвиваються ендоспорично; живуть за рахунок матеріалу, який був забезпечений спорофітною рослиною. Самець гаметофіт дуже недовговічний і має мало зберігається матеріалу (мікроспора невелика), але жіночий гаметофіт значно більший і живе місяцями на матеріалі, присутньому в спорі. Ранній ріст спорофіта, з жіночого гаметофіту, який присутній у мегаспорі, виглядає як проростаюче насіння (рис. 14), і ми побачимо в наступному розділі, що це аналогії - не вважається, що насіннєві рослини розвинулися з лікопогів. Ще однією особливістю деяких шипових мохів є те, що «жіночі» суперечки, ті, які переростають у жіночих гаметофітів, іноді розвиваються НА рослині спорофітів, ще одна особливість, яка повторюється у насіннєвих рослин.

