1.12: Грибкові статеві та грибкові групи
- Page ID
- 7191


«Вегетативна» (тобто не пов'язана з підлогою та розмноженням) структура більшості грибів, як правило, дуже послідовна: вони складаються з розгалужених ниток, які досліджують обсяг матеріалу, яким вони харчуються (будь то живий чи мертвий), і ця структура передбачає придбання матеріалів, які їм потрібні рости і виживати. Консистенція вегетативної форми ускладнює класифікацію грибів, оскільки вони здебільшого схожі, хоча деякі варіюються залежно від того, чи є нитки на коеноцитарних або септатних (з поперечними стінками і, отже, клітинними). Однак гриби виявляють варіацію, і їх можна групувати на основі структур, пов'язаних із статевим розмноженням. Винятками з цього є дві групи, де секс або повністю відсутній (гломеромікота - гриби, що утворюють ендомікори), або рідкісні (хітридомікота—хітриди). Решта три групи (хлібні цвілі = зигомікота, чашкові гриби = Ascomycota, і клубні гриби = Basidiomycota зазвичай проявляють специфічні особливості, пов'язані зі статевим розмноженням, які використовуються визначають ці групи (як правило, вважаються філа). Гриби, які не демонструють статевих ознак, раніше були введені в групу під назвою «Недосконалі гриби» (Deuteromycota), але молекулярні методи тепер дозволяють ці гриби бути поміщені в одну з груп, згаданих вище.
Гриби мають такі особливості, пов'язані з підлогою і розмноженням, які відрізняються від більшості звичних статевих організмів:
- R розмноження часто включає як сексуальні аспекти, так і безстатеві аспекти. Клітини, які називаються спорами - визначаються як клітини з як розсіювальними можливостями (тобто рухливими клітинами), так і репродуктивними можливостями (тобто клітини, які можуть перерости в нові грибкові організми), часто важливі як в безстатевому, так і в статевому розмноженні.
- G амети (клітини, які можуть зливатися з іншими гаметами) досить відрізняються від того, що більшість вважали б «типовими»; вони, як правило, є гіфальними клітинами (частиною грибкових ниток), які мають здатність зливатися з іншими гіфами. Залежно від групи, ці особливі гіфи можуть мати або не мати структурних особливостей, які б відрізняли їх від звичайних гіф.
- Сингамія часто включає в себе два кроки, розділені в часі і часто в просторі. Це є наслідком того, що злиття гіф, службовців гаметами, як правило, не слідує відразу злиття ядер. Клітинний синтез називається плазмогамією, а ядерний синтез називається каріогамією. Плазмогамія з подальшою затримкою каріогамії дозволяє грибам мати новий стан, стан дікаріону, де клітина має два ядра («dikaryon» означає «два ядра»), по одному від кожного з батьків. Цей стан часто увічнюється: клітина дікаріона ділиться, тоді як обидва ядра діляться, тим самим утворюючи нову клітину, яка також є дікаріоном. Цей процес може тривати, виробляючи кілька клітин дікаріона і гіфи дікаріона. Зверніть увагу, що хоча клітина дікаріона має дві копії кожної хромосоми, вона не вважається диплоїдною, оскільки кожне ядро є гаплоїдним, з лише однією копією кожної хромосоми.
- У якийсь момент деякі клітини дікаріона стають диплоїдними в результаті злиття двох ядер (каріогамія). Диплоїдні клітини НЕ увічнюються; вони піддаються мейозу з утворенням гаплоїдних ядер, Які розвивають стінки, щоб стати гаплоїдними клітинами. Таким чином, диплоїдний стан є дуже коротким за ступенем (тобто розміром і кількістю клітин) і часто коротким за часом, а також - диплоїдні клітини ніколи не діляться мітотично, утворюючи більше диплоїдних клітин, вони діляться лише мейотично, утворюючи гаплоїдні клітини. Ці гаплоїдні клітини є або незабаром стають спорами і диспергуються від батьківського гриба до нового місця, де вони проростають і утворюють гаплоїдні гіфи.
- «Спарювальні штами» - поширений механізм, якийгарантує, що гриби не спаровуються з собою. Всередині конкретного виду існує два-багато типів спарювання. Конкретний тип спарювання повинен знайти інший тип спаровування, щоб взаємодіяти статево (тобто, гіфи плавлення). Найпростіша ситуація має два типи спаровування: «+» і «-», і їм потрібно знайти один одного, щоб спаровуватися, але може бути більше двох типів, наприклад, типи A, B, C, D, E і A можуть спаровуватися з будь-яким з іншихАле не з іншим A. Взагалі взаємодія або його відсутність включає в себе хімічні сигнали (феромони) які виділяються одним типом спарювання і відчуваються тільки грибами іншого типу спарювання. У відповідь на феромон гіфи ростуть і знаходять один одного, викликаючи плазмогамію.
ТЕМИ
-
- Зигоміцети (формочки для хліба)
- Аскоміцети (чашечкові гриби)
- Базидіоміцети (клубні гриби)
Зигоміцети, форми для хліба


У зигоміцетів (рис. 2-3) більшість розмноження носить безстатевий характер і виникає в результаті вироблення стеблоподібної структури (спорангіофора), що закінчується кулястим спорангієм. Усередині спорангія є єдиною клітинною тканиною, що виробляється групою, тобто це структура, де окремі ядра упаковані один до клітини. Ці клітини дозрівають в суперечки, які розсіюються при розпаді спорангія. Набагато рідше відбувається розмноження, пов'язане зі статевим процесом і специфічною будовою, зигоспорою, від якої група і отримує свою назву. Процес ініціюється, коли дві сумісні гіфи відчувають присутність один одного через феромонів. Це запускає гіфи ростуть назустріч один одному і виробляють спеціалізовані гіфальні гілки, які здатні зливатися один з одним. Оскільки хлібні форми є коеноцитарними, злиття двох гіф (плазмогамія) дозволяє множинним ядрам вступати в близькість (рис. 4). Незвичайні для групи дві поперечні стінки, по одній в кожній з зростаючих гіф, утворюються поблизу точки злиття, створюючи єдину коеноцитарну клітину з ядрами, отриманими від кожного з двох спаровуються штамів (рис. 5-6). Це єдина клітина дікаріона, вироблена хлібними формами, і вона розвивається в структуру, звану зигоспорою (рис. 7-8), трохи збільшуючи і розвиваючи товсту стінку. Усередині зигоспор гаплоїдні ядра одного типу пара зядрами іншого типу і плавляться (каріогамія) утворюють диплоїдні ядра, перетворюючи клітину з дикаріона в диплоїдну, але вона все ж багатоядерна. Зигоспора, як правило, спить і, як правило, має більш специфічні вимоги до проростання та довший термін життя, ніж безстатеві спори. Коли зигоспора проростає, вона виробляє спорангіофор, порівнянний з тими, що виробляються безстатевим шляхом зі спорангієм на кінчику. У міру розвитку диплоїдні ядра піддаються мейозу, а гаплоїдні ядра індивідуально упаковуються в спори, коли виробляються клітинні стінки. Спори згодом диспергуються, і при проростанні утворюють гаплоїдні, коеноцитарні гіфи. Зверніть увагу, що стан dikaryon обмежений однією клітиною і має дуже коротке існування.

Аскомікота — чашечкові гриби

На відміну від хлібних форм, чашкові гриби (Ascomycetes) є перегородками, тобто вони мають поперечні стінки, а «живильні гіфи», ті, що набувають поживні речовини, побудовані з клітин, які мають єдине гаплоїдне ядро. Багато членів групи розмноження в першу чергу до виключно безстатевого, наслідком виробництва спеціалізованих гіфальних гілок, які виробляють дрібні клітини (конідії), які легко відламуються, щоб служити пропагулами. Як правило, ці клітини перебувають у стані спокою та мають інші клітинні особливості, наприклад, збережену їжу, які сприяють їх ролі у розмноженні та розгоні. Коли і якщо відбувається статеве розмноження, воно ініціюється, коли сумісні штами спаровування знаходяться близько один до одного і спілкуються феромонами. Виробляються роздуті гіфальні гілки, по одній на кожному штамі, продукуючи клітини, які трохи відрізняються за формою і є багатоядерними. Коли вони знаходяться близько один до одного, одна з клітин виробляє розширення, яке стосується розширеної клітини іншого типу спаровування і зливається з нею (відбувається плазмогамія) і гаплоїдні ядра переміщаються з однієї з структур, названих антеридієм і вважається чоловічим, до іншої, званої аскогонієм і вважається жіночим. В результаті міграції ядер клітина архегонії стає дікаріоном, з двома типами ядер, по одному від кожного з батьків. Аскогоній розвиває розширення і два ядра (по одному від кожного типу спаровування) асоціюються і мігрують в них. Утворюється поперечна стінка, яка виробляє першу клітинку того, що описується як аскогенна гіфа. Ці гіфи ростуть з кінчика і залишаються дікаріоном, з двома гаплоїдними ядрами на клітину, в результаті координації розширення кінчиків, двох ядерних відділів (по одному на кожне ядро), міграції ядра та утворення перегородки. Зростання аскогенних гіф плюс зростання обох (гаплоїдних) батьківських гіф виробляють плодове тіло густо переплетених гіф. Розмір плодового тіла коливається в величезних розмірах, приблизно від 100 мкм до 10 см і більше, причому більшість на меншому кінці. Розмір і форма варіюються і можуть бути використані для ідентифікації видів. Загальна назва групи ('cup fungi') відноситься до плодового тіла у формі чашки. Чашкові гриби зустрічаються як живуть самостійно, так і як грибковий партнер ряду лишайників; багато лишайників виробляють аскокарпи (рис. 10), які відносно великі і помітні неозброєним оком. Інші плодові тіла мають колбоподібну форму або повністю закриті і зазвичай мають протяжність менше мм.

У конкретній частині цього плодового тіла статевий цикл завершується. Кінчики аскогенних гіф утворюють гачок, повертаючись назад на себе і два ядра зливаються (відбувається каріогамія) в осередку, яка утворила гачок (крозье), роблячи її (коротко) диплоїдної клітиною. Єдині диплоїдні клітини, знайдені в Ascomycota, - це ці специфічні клітини, зрештою називаються asci (сингулярні = ascus), розташовані на кінцях аскогенних гіф (ці гіфи не виробляють додаткових клітин після формування гачка). Потім диплоїдна клітина подовжується і піддається мейозу, утворюючи подовжену клітину з чотирма гаплоїдними ядрами. Кожен з них потім проходить через мітоз, щоб виробляти подовжену клітину, яка називається аскусом з вісьмома гаплоїдними ядрами поспіль. Кожне ядро набуває клітинну стінку і переростає в те, що називається аскоспорою. Asci з вісьмома аскоспорами є діагностичними для Аскомікота.

Гідростатичний тиск всередині аска викликає примусове розсіювання аскоспор при розриві верхівки мускуса. Розсіяні аскоспори проростають, утворюючи гаплоїдні гіфи, які утворюють гаплоїдні міцелії, які можуть розмножуватися безстатевим шляхом через конідії або інші спори. Якщо гаплоїдний міцелій контактує з гіфами іншого типу спарювання, може бути спровоковано статеве розмноження.


Базидіоміцети — клубні гриби
Клубні гриби є перегородками, як чашкові гриби (Ascomycota). З усіх грибкових груп клубні гриби мають найбільш великий дікаріонний стан. Як і у випадку майже у всіх грибів, стан дікаріона ініціюється, коли два сумісних гаплоїдних, монокаріонних гіфи знаходять один одного в результаті хімічного тяжіння. Плазмогамія відбувається, утворюючи клітину дікаріона. Ця клітина широко ділиться і росте, утворюючи живильний міцелій, який є дікаріоном. Багато з базидіомікоти виробляють «затискні з'єднання» (рис. 17), які можуть допомогти зберегти стан дікаріона, оскільки гіфи подовжуються. Ядра піддаються мітозу, і утворюються поперечні стінки. У міру подовження кінцевої клітини між кінцевою клітиною і її батьківською клітиною утворюється дуга. Коли два ядра діляться, дуга дозволяє одному з двох ядер переміщатися до батьківської клітини у вигляді перегородки. Це гарантує, що і дочірня клітина, і материнська клітина мають по одному з кожного типу ядра.
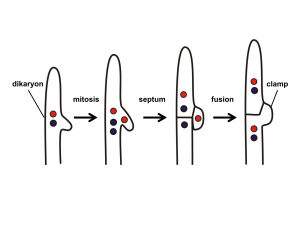
Нагадаємо, що в хлібних формочках (Zygomycota) є лише одна клітина дікаріона і єдина дікаріонна клітина грибків чашечки (Ascomycota) знаходяться в плодових тілах. Таким чином, якщо хтось стикається з дикаріотичними гіфами поза плодовим тілом, то він повинен належати клубному грибу (Basidiomycota). Дикаріотичні гіфи клубних грибів ростуть і харчуються інтенсивно, поки не будуть виявлені відповідні умови, щоб спровокувати «плодоношення». У цей момент картина росту хоча б деяких гіф змінюється від тієї, де досліджується їх субстрат для харчування, до тієї, де гіфи переплітаються між собою, утворюючи щільну масу гіф, які стануть плодовим тілом. Часто плодове тіло виходить з субстрату, яким харчується гриб, наприклад, гриб, що виходить з грунту або зі стовбура дерева. Десь на або в щільній масі гіф утворюються спеціальні клітини, звані базидіями, які мають «клубовидную форму» (рис. 18). Усередині цих клітин відбувається каріогамія, перетворюючи їх вдиплоїдні клітини. Диплоїдне ядро піддається мейозу, утворюючи чотири гаплоїдні ядра, але цитокінезу не відбувається, що робить базидій «квадра-нуклеатом». Чотири розширення виростають з базидію, і чотири ядра мігрують в них. Далі слід формування клітинної стінки для отримання базидію без ядра, але з чотирма слабо прикріпленими гаплоїдними клітинами, званими базидіоспорами, що відходять від них. Спори балістично виділяються і розганяються вітром. Коли вони проростають, вони утворюють нові гаплоїдні гіфи.

На відміну від чашкових грибів, плодові тіла яких, як правило, невеликі і зазвичай не помічені, плодові тіла багатьох клубних грибів часто відносно великі, 10-30 см і мають характерні форми, які ми описуємо як гриби (зі стеблом і капелюшком) (рис. 19-20) або брекет-гриби ( грубо копитної форми і прикріплюється без стебла до стовбурів деревних рослин). Двома дуже поширеними схемами розподілу базидій є (1), що поширюються від тонких плавників тканини під назвою «зябра», типових для комерційно доступного гриба, або (2) базидії, що виходять з трубок (циліндрів) тканини, які, якщо дивитися на поверхню, виглядають як численні пори.





Дві великі групи базидіоміцетів, сажки (рис. 22-23) (клас устоміцетів) та іржі (рис. 24-25) (клас теліоміцетів) мають дещо різні, а іноді залучені моделі статі та розмноження, пов'язані з їх облігатним паразитичним способом життя. Багато з цих організмів дуже важливі економічно, оскільки вони можуть різко знизити врожайність важливих культур. Ні утворюють «плодові тіла», виготовлені виключно з грибкового матеріалу, а скоріше викликають аномальний ріст (галли) на рослині, в якій вони ростуть. Ці галли складаються з заражених рослинних клітин, незаражених клітин рослин, що ростуть аномально через паразита, і деяких грибкових гіф, що з'єднують заражені клітини. Хоча описаний вище типовий базидіоміцет має один тип спор, гаплоїдні базидіоспори, що утворюються в плодовому тілі, як іржа, так і сажа зазвичай виробляють теліоспори, від однієї до декількох клітинних спор, які містять дікаріотичні клітини. При проростанні теліоспор відбувається каріогамія з подальшим мейозом, продукуючи базидій з гаплоїдними ядрами, які виробляють гаплоїдні базидіоспори.

Хоча більшість сажки виробляють лише ці два типи суперечок, іржа може мати ще два-три типи суперечок та складний життєвий цикл, який іноді включає двох господарів. Життєвий цикл пшеничної іржі показаний на малюнку 26 і розглянутий в главі 30.

Базидіоспори заражають господаря #1 (барбарис) і утворюють структуру під назвою пікнії на верхній поверхні листа. Pycnia виробляють дві структури, які спричиняють сингамію. Однією з них є гаплоїдні спори, які називаються пінкоспорами (або сперматією), які можна вважати гаметами. Пікноспори виділяються як поодинокі клітини в солодкий «нектар», який приваблює комах, які можуть транспортувати їх до інших сумісних (тобто різного типу спарювання) пікнії, де вони можуть зливатися (плазмогамія) з другою структурою, виробленою пікнією, що беруть участь у сингамії: «гнучкі гіфи», що виходять з пікнії. Злиття пінкоспори з гаплоїдними клітинами цих гіф спричиняє сингамію і утворює початкову клітину гіфи дикаріона, яка росте на дно листа, утворює кульку і вивільняє спори дикаріону (ацеоспори), які заражають господаря #2 (пшениця), змушуючи його утворювати інший тип спори дикаріону, урідиноспори, які можуть заразити більше рослин пшениці. В кінці сезону на пшениці утворюється кінцевий вид спор - теліоспора. Каріогамія і мейоз виникають у теліоспор і з них виходить промицелія (в основному базидії) і виробляють базидіоспори.
