7.2: Тьюрингоподібна модель для генерації смуг у розробці цифр (Рівера та Рамірес)




- Last updated
- Oct 24, 2022
- Save as PDF

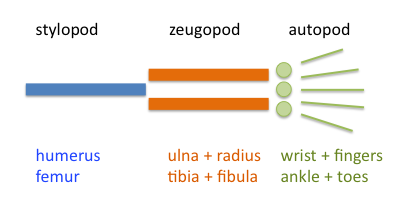
Ваші кінцівки спочатку починають рости як невеликий нубін від вашого тулуба. У міру розмноження клітин невеликий стрижень скелетної тканини починає рости прямо посередині. Це стане плечовою кісткою або стегновою кісткою. Ця скелетна тканина визначається на початку Sox9, фактор транскрипції також бере участь у визначенні статі. У міру зростання тканини кінцівки довше і ширше з'являються дві паралельні кістки (променева і ліктьова або великогомілкова і малогомілкова кістка). Тканина все ще росте і ширше, і з'являються п'ять паралельних кісток - п'ясткових і в кінцевому підсумку фаланги (рис. 3).
 |
 |
Малюнок 3: Частини чотириногої кінцівки. Ліва панель — це зображення з Вікісховища (Peteruetz), опубліковане під ліцензією CC BY-SA 3.0. На правій панелі показана базова модель розвитку чотириногих кінцівок. У міру утворення кожного хряща ущільнення (синій, Sox9) він стає репресивним до утворення довколишніх ущільнень. Дифузія і сила репресії дають «довжину хвилі», показану фіолетовим кольором, поза хвилею можуть утворюватися конденсації (вісь X - проксимально-дистальна вісь, а вісь Y - сила репресії). Жовта область - область нестабільності (висока концентрація ФГФ з верхівкового ектодермального хребта), де конденсації не можуть утворюватися. У міру того, як кінцівка зростає, є місце для більшої кількості конденсатів з однаковою довжиною хвилі. Редагований файл svg правої панелі можна завантажити за адресою https://scholarlycommons.pacific.edu/open-images/32/
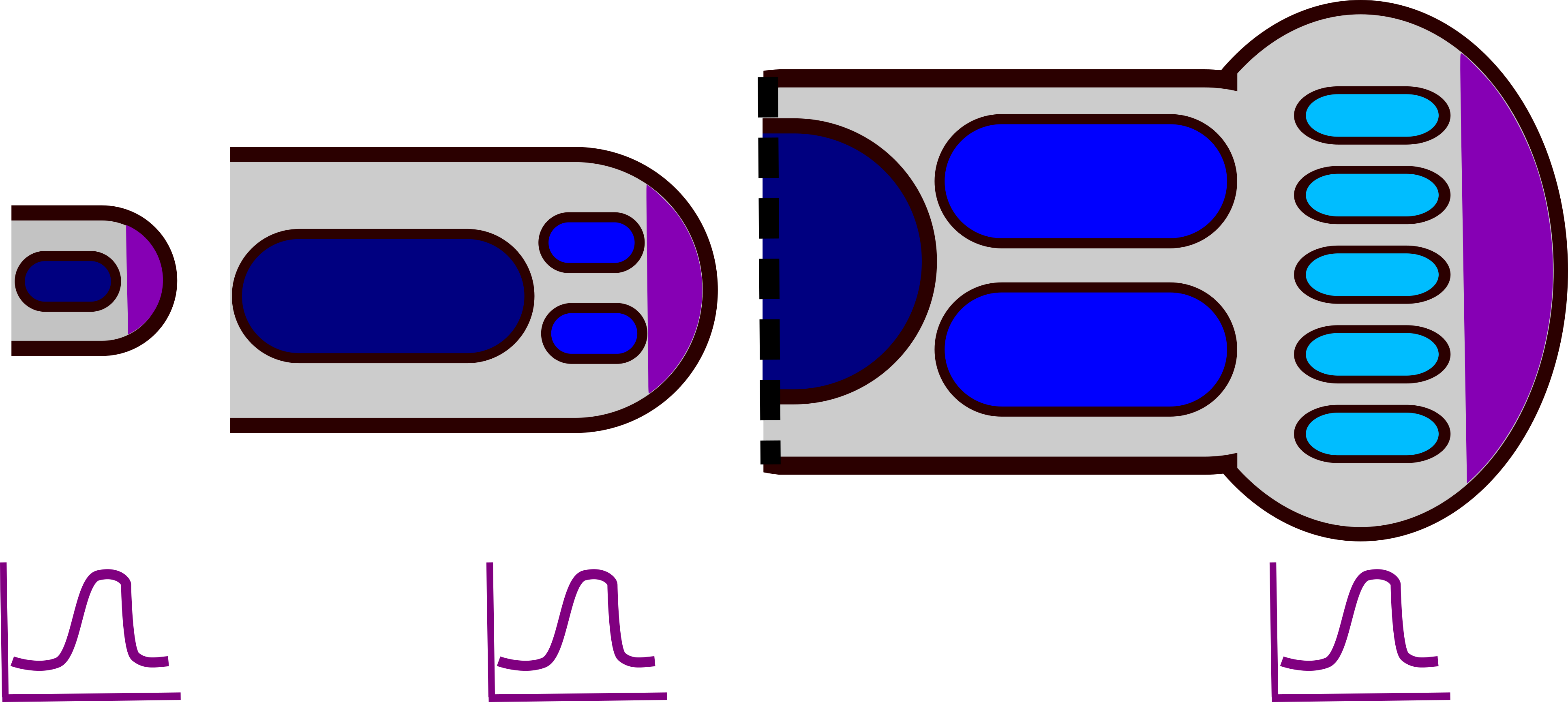
Рання модель розвитку кінцівок припускала, що інгібуючий дифузний морфоген виробляється конденсаціями хряща (які згодом стануть кістками кінцівок). У ранній нирці кінцівки цей інгібітор може дифузно поширюватися по всій крихітній тканині, тому закладається лише одна конденсація, утворюючи велику кістку стилопода. Оскільки кінцівка росте довше і ширше, інгібітор не може досягти достатньо високих концентрацій, щоб покрити всю ширину бруньки кінцівки, і утворюються дві ущільнення (паралельні кістки зеугопод). У розвивається аутопод тканина досить широка, щоб підтримувати 5 взаємно-гальмівних ущільнень - п'ять цифр стопи або кисті (рис. 3) 16. Кілька моделей Тьюринга можуть генерувати зразок смужки, включаючи модель інгібітора активатора та модель виснаження субстрату (рис. 4).
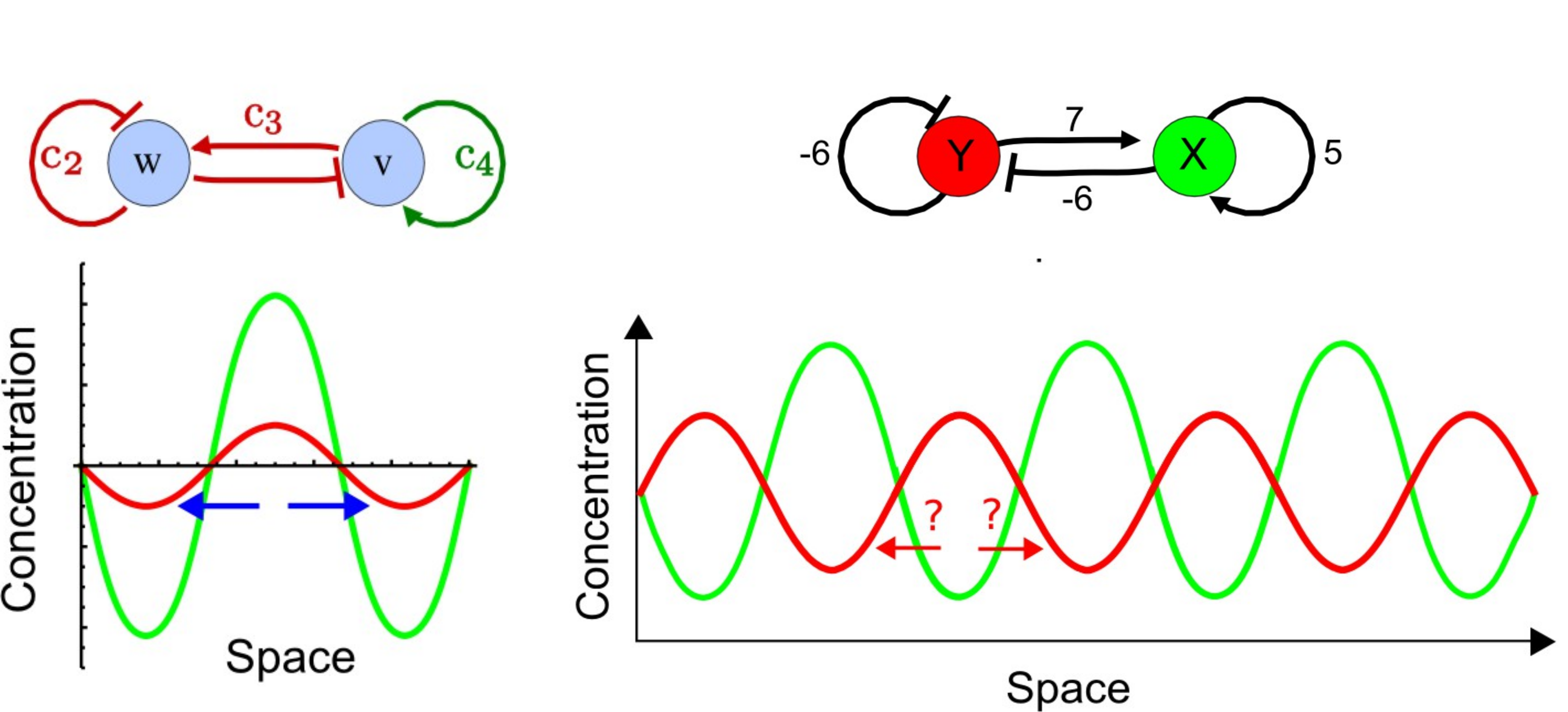 |
Малюнок 4: Дві моделі Тьюринга для розрядних смуг. Модель зліва - це модель інгібітора активатора, де дифузний активатор V активує себе, а також дифузний інгібітор W. Інгібітор W інгібітор W інгібує себе і активатор V. cs представляють різні сили взаємодії. Модель праворуч - це модель виснаження субстрату, де дифузний активатор X використовує (або інгібує) субстрат Y. Таким чином субстрат Y гальмує себе, активуючи власний інгібітор. Для тестування між цими моделями можна оцінити експресію молекул, які, як вважають, беруть участь у процесі. Якщо шаблони виразів перекриваються (графік ліворуч), то підтримується модель інгібітора активатора. Якщо шаблони виразів взаємодоповнюють (графік праворуч), то підтримується модель виснаження субстрату. Малюнок модифікований з оригінальних фігур Marcon et al, 2016 21, спочатку опублікований під ліцензією CC BY 4.0 |
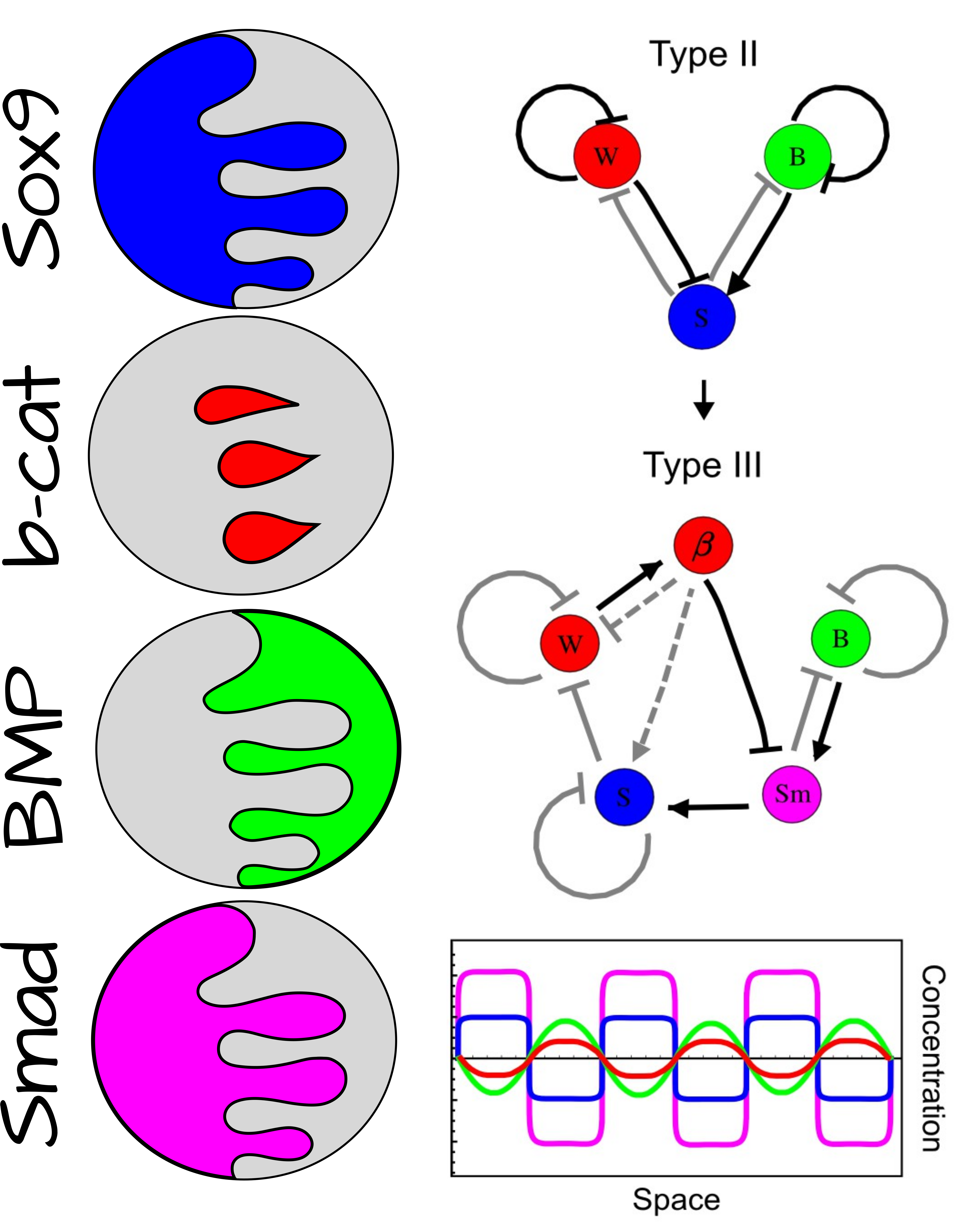
Експерименти над білками, вираженими або в міжцифрових областях, або в цифрах, забезпечили підтримку моделі виснаження субстрату 17. Однак ці експерименти також показали, що замість одиночних білків цифрове малювання фактично визначається за допомогою декількох взаємодіючих сигнальних шляхів. Bmp, Sox9 та Wnt утворюють основні компоненти малюнка, що генерує «довжину хвилі» цифрової відстані. Всі три з цих генів утворюють градієнти експресії, з найвищим Sox9 посередині цифр і найвищим Bmp і Wnt в міжрозрядних (рис. 5) 17. Bmp, виражений в міжрозрядних областях, пригнічує себе, але дифундує назовні, щоб активувати Sox9 в сусідніх осередках (майбутні цифри). Sox9 пригнічує Bmp, гарантуючи, що Bmp виражається лише в міжцифрах. Ліганди Wnt повсюдно поширені, але його цілі (включаючи b-катенін) виражаються лише в міжцифрах. Втрата b-катеніну призводить до розширення експресії Sox9, а розширення експресії b-катеніну зменшує експресію Sox9, припускаючи, що WNT/B-катенін є репресором Sox9 19,20. Ці взаємодії дають модель, де цифри утворюються в регулярній схемі з довжиною хвилі, визначеною силою взаємодій Bmp/Sox9/Wnt і здатністю Bmp і Wnt до дифузії (рис. 5) 17.
 |
|
|
Малюнок 5: Распопович та ін. 17 модель для специфікації цифр. Sox9, первинний цифровий маркер, вказує клітини для формування скелетних ущільнень. Вона виражається в осередках, які стануть цифрами в автоподе. У цій моделі Sox9 (синій) є активатором і він взаємодіє з сигнальними шляхами BMP і Wnt. Автоподи відображаються у вигляді сірих кіл у лівій колонці з кольоровими візерунками виразів. Sox9 і Smad (фіолетовий, вихід шляху BMP) виражаються цифрами, тоді як BMP (зелений) та b-катенін (червоний, вихід шляху Wnt) виражаються в міжцифрах. У лівій колонці показана модель Тьюринга для цих взаємодій. Топ-модель спочатку була запропонована Raspopovic et al 17, а згодом вдосконалена Marcon et al 21. Модель Marcon et al підтримує центральну роль Sox9, що пригнічує власне вираження поза конденсацією. Sox 9 пригнічує шлях Wnt, який пригнічує b-катенін та інші гені-мішені в зонах Sox9. b-катенін пригнічує Smad, який є мішенню дифузної сигналізації BMP. Інгібуюча дія Smad на BMP гарантує, що BMP не активний у SMAD-експресуючих клітинам, надаючи цим двом членам шляху BMP взаємодоповнюючі схеми активації. У нижній правій панелі підсумовується висновок складної моделі Marcon et al. Змінено з Marcon et al 21, спочатку опубліковані під ліцензією CC BY 4.0 Редаговану версію svg цього малюнка можна завантажити за адресою: https://scholarlycommons.pacific.edu/open-images/33/ |
Малюнок 6: Зміна довжини хвилі. Автопод у формі весла повинен забезпечувати додаткове утворення конденсату в міру зростання. Це повинно привести до додаткових кісток в дистальному відділі автопода або розгалуження наявних ущільнень (верхня панель). Мутанти Hoxd13 дають фенотипи, такі як верхня панель, припускаючи, що Hoxd13 змінює довжину хвилі конденсацій Sox9. Hoxd13 дифундує від верхівкового екодермального хребта і утворює градієнт (жовтий). Там, де рівні Hoxd13 високі, довжина хвилі (або міжзначний простір) велика. Там, де рівні Hoxd13 низькі, довжина хвилі невелика. Модуляція рівнів Hoxd13 та/або його здатність регулювати довжину хвилі призводить до нерозгалужених цифр 17,18. Редаговану версію svg цього малюнка можна завантажити за адресою: https://scholarlycommons.pacific.edu/open-images/34/ |
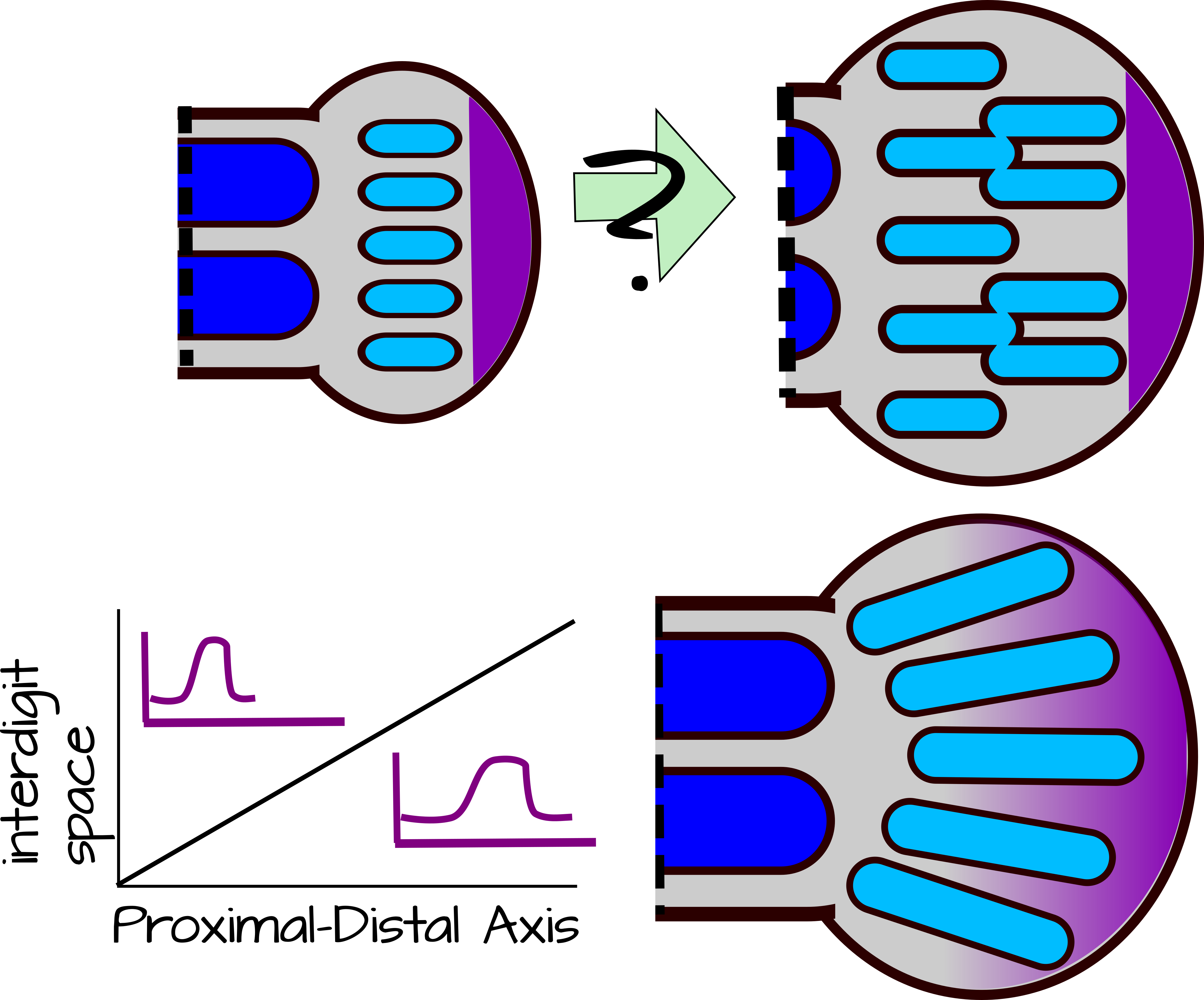
Але є одна велика проблема з цим. На відміну від зеугопод і стилопод, автопод має веслоподібну форму. Це означає, що він швидко розростається під час розвитку, утворюючи тканину, яка повинна підтримувати більше ущільнень хряща на кінчику, ніж біля зап'ястя запропонованою моделлю (рис. 6). Цю проблему вирішує Hoxd13. Hoxd13 - це ген Hox, який допомагає малювати руки, ноги, сечовивідні шляхи та репродуктивні органи. У руках і ногах (автоподів) це виражається в градієнті, з найвищими рівнями біля верхівкового екодермального хребта (AER) на кінчику бруньки кінцівки. Hoxd13 стабілізує цифрове число двома способами. По-перше, він діє як «зона без малюнка». У регіоні з найвищим Hoxd13 вираз Sox9 нестабільний і нові конденсації не можуть утворюватися. Далі від AER в районі нижнього Hoxd13 можуть утворюватися нові конденсації, керовані SOX9. Ще далі від AER, де немає виразу Hoxd13, існуючі конденсації стабільні. Ніяких нових конденсатів не утворюватиметься навіть у міру того, як тканина розростається ширше. Hoxd13 також стабілізує цифрове число за рахунок збільшення здатності Sox9 інгібувати Bmp і Wnt. Це призводить до того, що довжина хвилі цифри ширше в дистальній частині автопода, пригнічуючи розвиток додаткових цифр там (рис. 6) 18. У мишачих мутантів Hoxd13 хрящові ущільнення гілки автопода в дистальних ділянках, слідуючи природній довжині хвилі взаємодії Bmp/Sox9/Wnt 18.
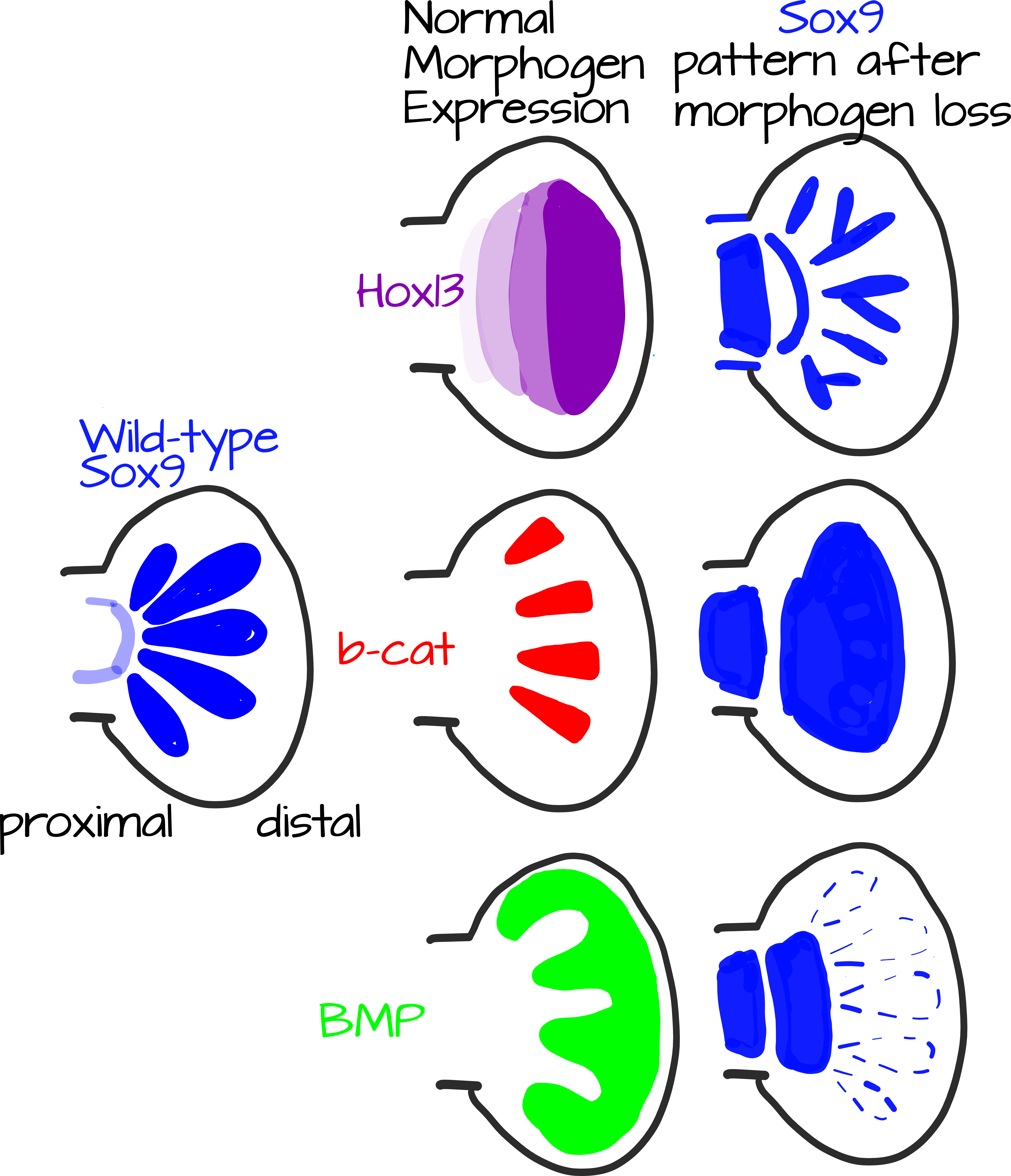 |
Малюнок 7: Мутації в регуляторах вираження Sox9. Sox9 в нормі виражається в хребетних скелетних елементах (майбутніх кістках і хрящах). Зліва - типовий шаблон вираження Sox9 в автоподі. Середній стовпець показує нормальну експресію трьох морфогенів, що беруть участь у малюнку Sox9 в автоподі. Hox13 (фіолетовий) походить від AER і утворює градієнт, який вище на дистальному кінці і нижче на проксимальному кінці. b-cat (бета-катенін, червоний) є найвищим між цифрами (в міжрозрядних). БМП (зелений) є найвищим всюди, крім скелетних елементів, в тому числі і в міжпальцевих. Втрата функціональних мутацій у кожному з цих генів виявляє їх функцію в налаштуванні механізму Тьюринга для формування цифр. Hox13 зазвичай встановлює довшу довжину хвилі Sox9 в дистальному кінці автопода. Мутанти Hox13 демонструють проксимальну довжину хвилі на дистальному кінці - в результаті чого розгалужуються цифри. b-cat зазвичай запобігає вираженню Sox9 в міжрозрядних. B-кішки мутанти демонструють скелетні елементи через аутопод. BMP в нормі викликає секрецію Smads, які викликають експресію Sox9 в клітині, що не експресують BMP. БМП мутанти призводять до втрати скелетних елементів. Фігура Десмонда Раміреса |
 |
Д-р Десмонд Рамірес, доктор філософії є докторантом, який працює в лабораторії Кац в UMass, Амхерст. Дізнатися більше про його роботу можна на сайті https://desmondramirez.wordpress.com/ |