1.4: Основні метаболічні шляхи
- Page ID
- 26068

Основний огляд метаболізму
- Гліколіз (цитозол) і цикл ТСА (мітохондрії) перетворюють глюкозу в молекули високої енергії: ________ і ___________ і __________.
Це тільки початок виробництва енергії. NADH і FADH 2 можуть бути перетворені в більше ATP. Окислювальне фосфорилювання - це метаболічний шлях, який передає енергію від НАДГ до синтезу АТФ в мітохондріях.
- Окислення NADH відбувається протягом багатьох етапів. Чому клітини не роблять цю реакцію безпосередньо? (Підказка: Це реакція гідридів!)
Місця стільникового зв'язку
Електрони, що зберігаються у вигляді відновлених коферментів, NADH або FADH 2, пропускають через ланцюжок білків і коферментів для зменшення O 2 - кінцевого акцептора електронів - в H 2 O.
- НАДГ утворюється в який момент метаболізму: ___________.
- Цикл ТСА відбувається в __________.
- Цей електронний перенесення окислювального фосфорилювання відбувається в ________________.
Виробництво АТФ
Енергія, що виділяється електронами, що протікають через цей ланцюг транспорту електронів, використовується для транспортування протонів для генерації градієнта рН через мембрану.
- Фосфорилювання АДФ з утворенням АТФ є [ендотермічним або екзотермічним].
- Протони стікають назад через мембрану для відновлення рівноваги. Цей процес є [дифузією або активним транспортом] і може керувати реакцією.
Основний метаболізм: гліколіз
Глюкоза метаболізується для отримання енергії (АТФ) для клітини з вивільненням CO 2 і H 2 O в якості побічних продуктів. Гліколіз - це ряд ферментно-каталізованих реакцій, які розщеплюють глюкозу на 2 еквіваленти пірувату. Цей процес (узагальнено нижче) також називають стежкою Емден-Мейєргофа.
- Скільки АТФ виробляється в цьому процесі? Майте на увазі, що все подвоюється після того, як глюкоза 6 С розщеплюється на одиниці 2 3С.
- Скільки споживається АТФ?
- Гліколіз призводить до чистого утворення:
- ______ НАД
- ______ АТП
- ______ Ч 2 О
- Гліколіз - це процес в гору або під гору? (Тобто екзотермічний або ендотермічний?)
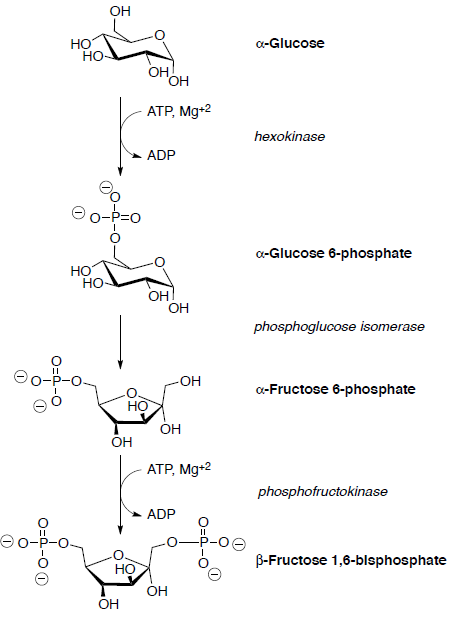
Припустимо, що всі реакції відбуваються всередині ферменту.
Глюкозу спочатку фосфорилюють в гідроксильній групі на С6 шляхом реакції з АТФ.
- Запропонуйте механізм цієї реакції.
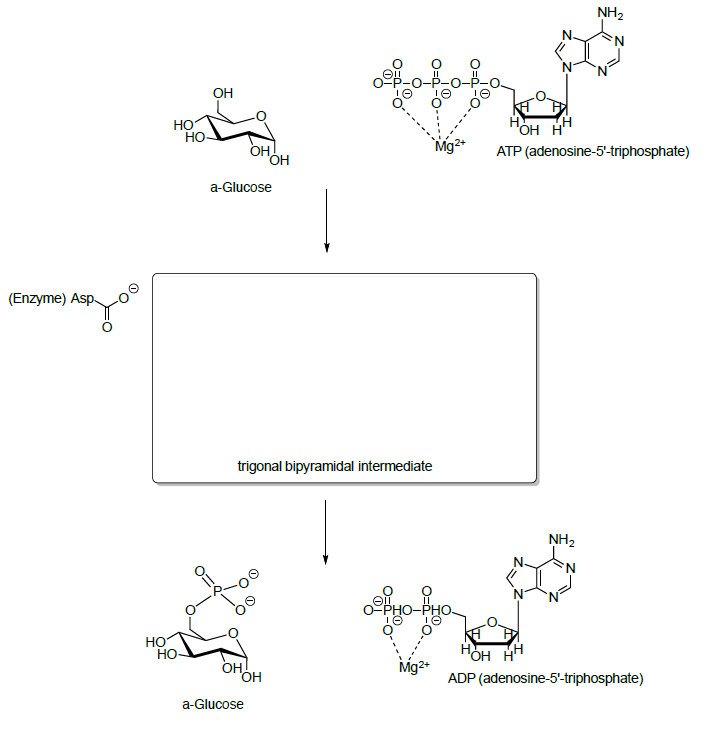
- АТФ не є тим, що реактивний сам по собі. Чому?
- Поясніть, чому фосфатний ефір є хорошим електрофілом, коли Mg +2 знаходиться навколо.
Глюкозо-6-фосфат ізомеризується до фруктозо-6-фосфату на наступному етапі. Взаємоперетворення глюкози - фруктоза - це багатоступінчастий процес, деталі якого ще не до кінця вивчені.
Починається з розкриття геміацеталя до альдегіду з відкритим ланцюгом.
- Запропонуйте механізм цієї реакції.
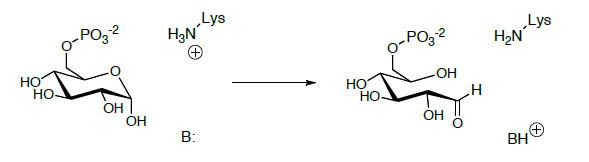
Альдегід з відкритим ланцюгом піддається кето-енолової тавтомеризації до ендіолу, який додатково таутомеризується до іншої кето-форми.
- Дивлячись на структури цукрів, запропонуйте механізм цієї реакції.
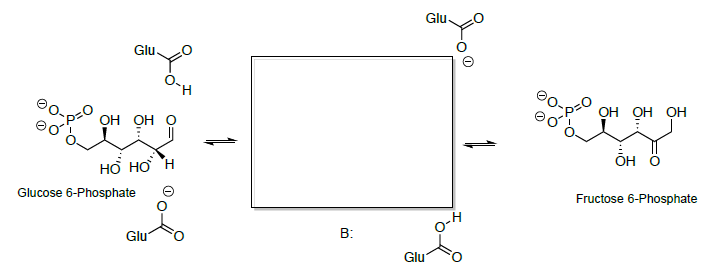
Циклізація гідроксикетона з відкритим ланцюгом дає фруктоза (геміацеталь).
- Show a mechanism.
- Predict the product.

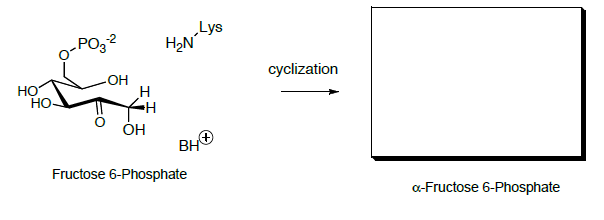
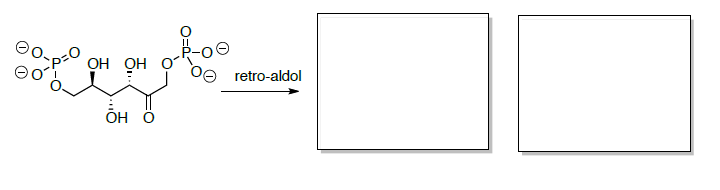
Потім фруктозо-6-фосфат перетворюється на фруктозу 1,6-бісфосфат, який згодом розщеплюється на два тривуглецевих сполуки через ретро-альдол.
Огляд: реакція альдола
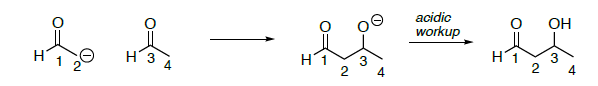
- На реакцію альдола вище,
- Покладіть коло навколо нуклеофіла
- Покладіть коробку навколо електрофілу у вихідних матеріалах
- Виділіть зв'язок, яка утворюється (порушена в ретро-реакції)
Ретро-Альдол
Якщо реакція підганяється на вихідні матеріали (ретро-альдол), то реакція буде сприяти вихідним матеріалам.
- Намалюйте механізм ретро-альдола при запуску фруктози 1,6-бісфосфату.
- Передбачте два продукти цієї ретро-альдольної реакції.
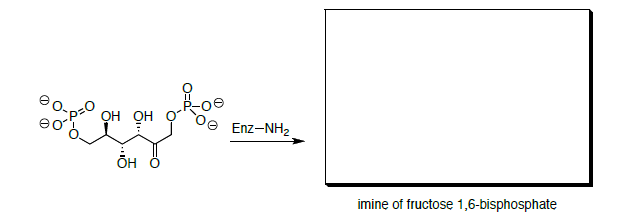
Цей механізм фактично комплектується іміном. Фруктоза 1,6-бісфосфат спочатку вступає в реакцію з аміногрупою залишку лізину з ферменту.
- Намалюйте механізм формування іміна.
Потім імін може зробити реакцію «ретро-лелеки емалі» (подібно до ретро-альдола).
Огляд: Лелека Енамін (адол з енаміном, що замінює аніон енолат як нуклеофіл).
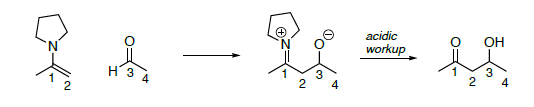
- На реакцію енаміну вище,
- Покладіть коло навколо нуклеофіла
- Покладіть коробку навколо електрофілу у вихідних матеріалах
- Виділіть зв'язок, яка утворюється (порушена в ретро-реакції)
Ретро-лелека енамін
Якщо реакція підганяється на вихідні матеріали (ретро-лелека енамін), то реакція сприятиме енаміну і альдолу вихідних матеріалів.
- Передбачте два утворилися продукти.
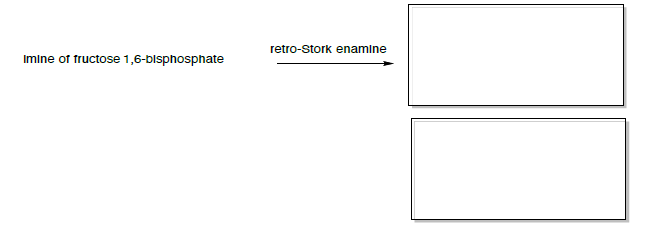
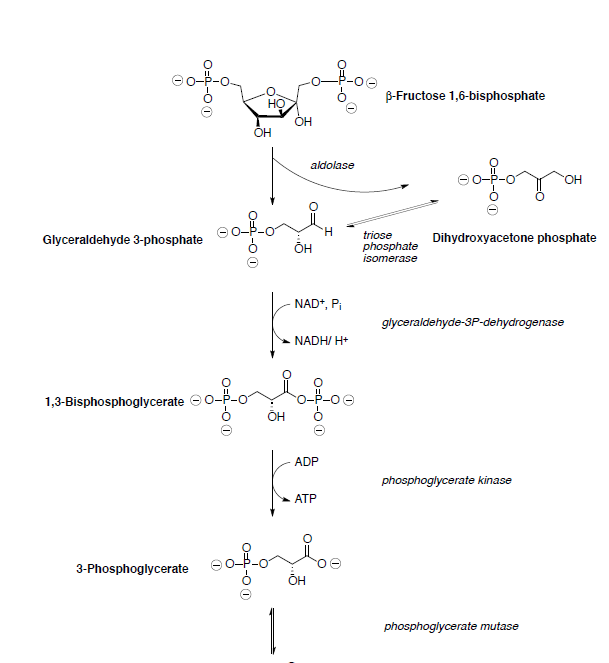
Продукцією енаміну ретро-лелеки є енамін дигідроксіацетону фосфату і гліцеральдегіду 3-фосфату (показано нижче).
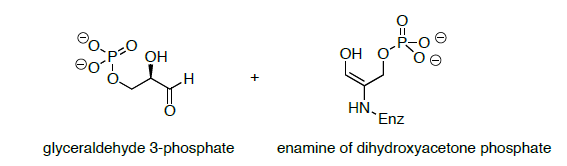
- Propose a mechanism for the conversion of the enamine of dihydroxyacetone phosphate is converted to a second molecule of glyceraldehyde 3-phosphate.
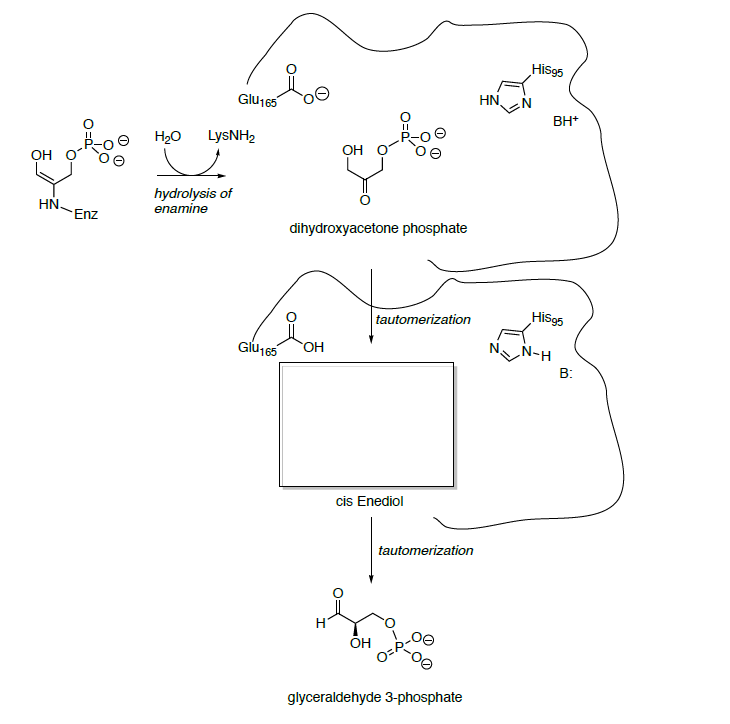
Glyceraldehyde 3-phosphate is oxidized and phosphorylated to 1,3-bisphosphoglycerate.
- Show the mechanisms for this transformation.
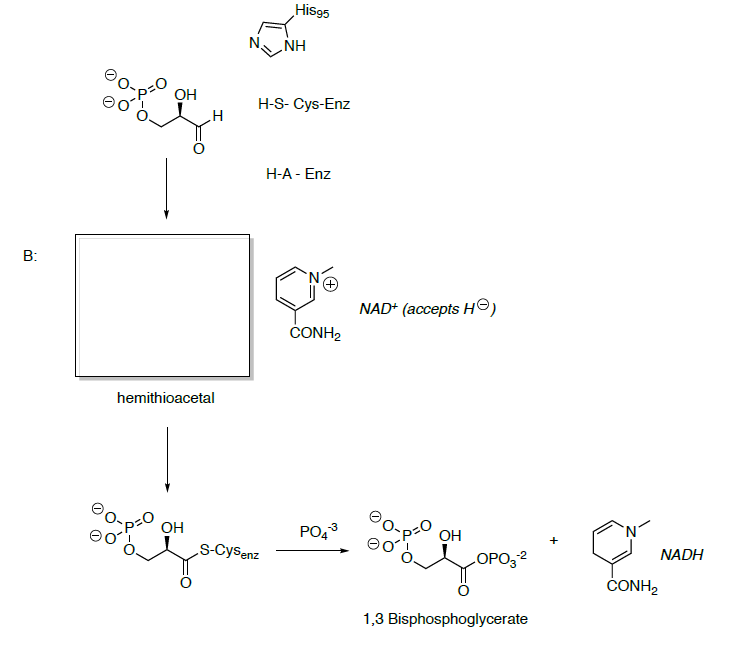
- What is the functional group formed in 1,3-bisphosphoglycerate?
- Predict the reactivity of this carbonyl.
Phosphoglycerate kinase catalyzes the transfer of a phosphoryl group from 1,3-bisphosphoglycerate to ADP forming ATP and 3-phosphoglycerate.
- Propose a mechanism for this transformation.
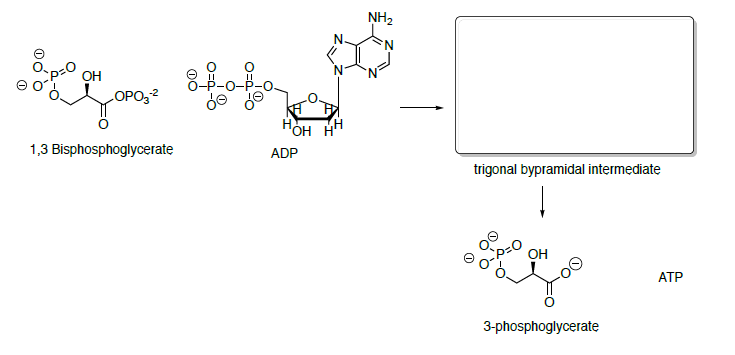
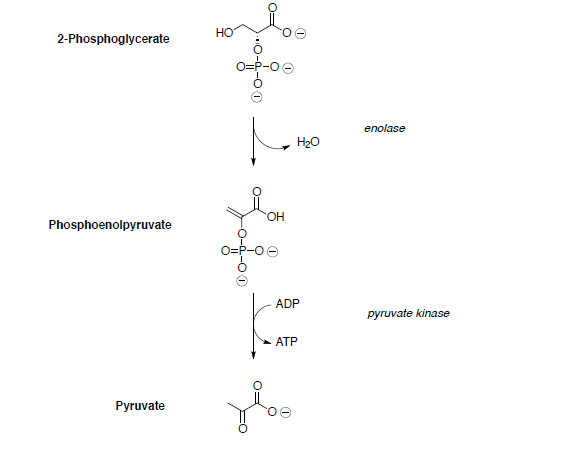
3-phosphoglycerate is converted to phosphoenol pyruvate (PEP) through dehydration and dephosphorylation.
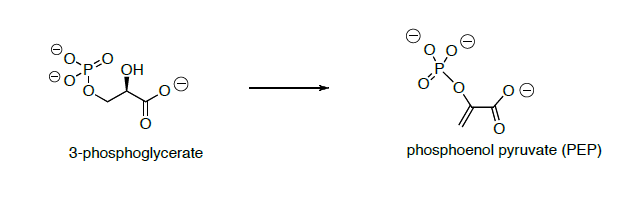
In the last step of the metabolic breakdown of sugars (glycolysis), an enol phosphate is converted to pyruvic acid (shown below). The pyruvic acid is then converted to Acetyl Co A, which is the beginning of the TCA cycle.
- Draw a mechanism for the conversion of the enol phosphate to pyruvic acid.
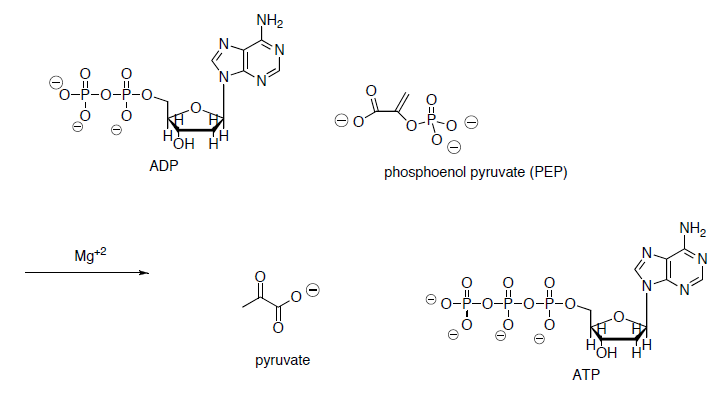
- What drives this reaction? (ie what factors make this reaction energetically favorable?)
Basic Metabolism: TCA Cycle
Hans Krebs and Fritz Lipmann shared the Nobel Prize for Physiology and Medicine in 1953 for their work on elucidating the Krebs cycle and coenzyme A. The Krebs Cycle [or tricarboxylic acid (TCA) or citric acid cycle] plays a central role in the metabolism of glucose to produce energy (ATP). The TCA cycle results ultimately in the oxidation of acetic acid to two molecules of carbon dioxide.
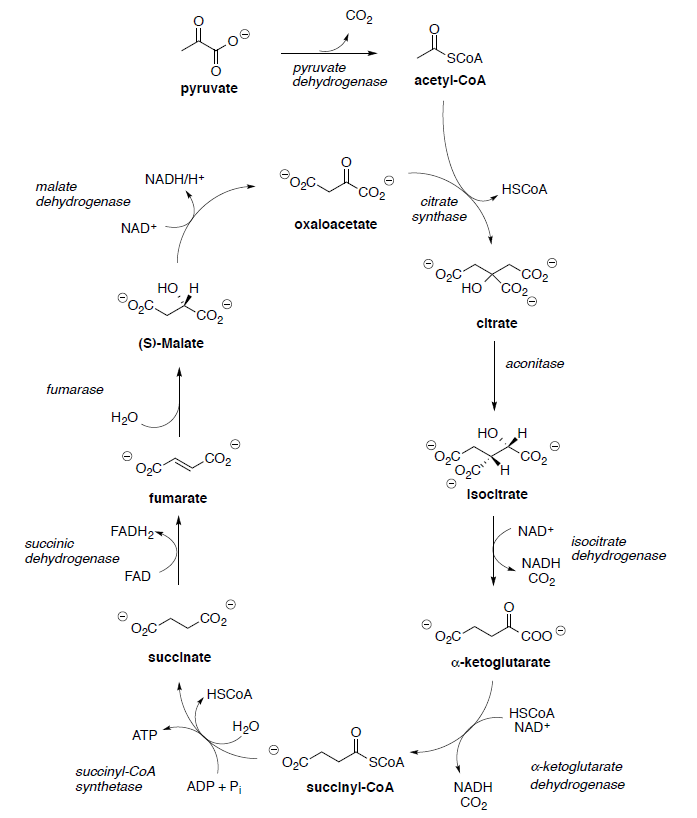
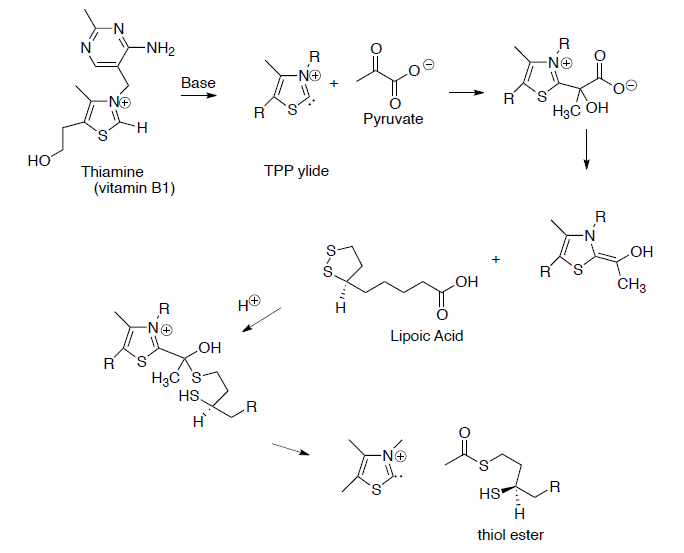
Pyruvate (end product of glycolysis) must be converted to acetyl CoA to enter the TCA cycle.
This process begins with the formation of a thiol ester from pyruvate.
- Draw reaction mechanisms for the steps shown below.
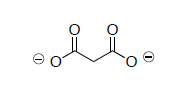
- In his experiments that led to the elucidation of the TCA cycle, Hans Krebs added malonate (shown below) to extracts of pigeon flight muscle. The malonate could not be used as a substrate to replace pyruvate in the pathway above. Why can’t malonate be used? (Think of the carbonyl hill).
At this point, co-enzyme A reacts with the thiol ester (formed in question on previous page) to form acetyl CoA (shown below). To help keep track of the sulfurs, one is in a box and one is in a circle.
- Draw the mechanism for this reaction.
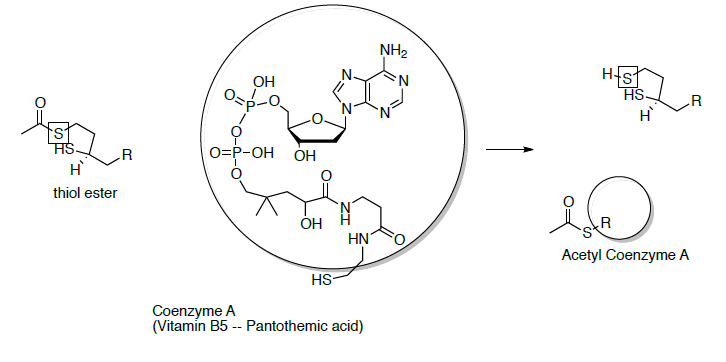
- The thiol ester formed in the last step of the reaction above is an ‘activated carbonyl’ (i.e. a better electrophile). Explain why the thiol ester is a better electrophile than the carboxylate anion.
- In an equivalent organic chemistry reaction, what would you use as an ‘activated carbonyl’?
In the next step Acetyl CoA reacts with oxaloacetate to form citryl CoA.
- Propose a mechanism for this reaction.
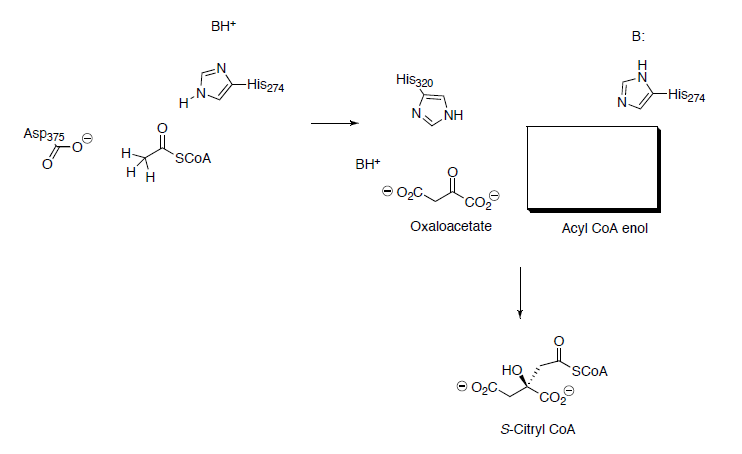
- In a similar reaction in organic chemistry, what would be the product for the reaction below? What type of reaction is this?
Citryl CoA is then hydrolyzed to citrate.
- Propose a reaction mechanism for this reaction.
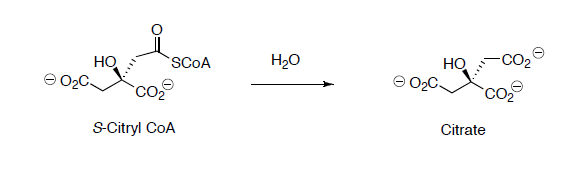
Citrate is converted to isocitrate through two steps.
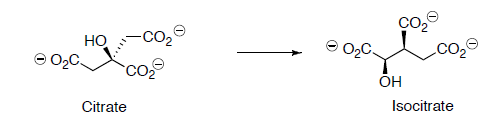
- Label all chiral centers with R or S.
- What changed in the conversion of citrate to isocitrate?
Isocitrate is oxidized to oxalosuccinate with NAD+.
- Draw the mechanism (and the other product) for this reaction.
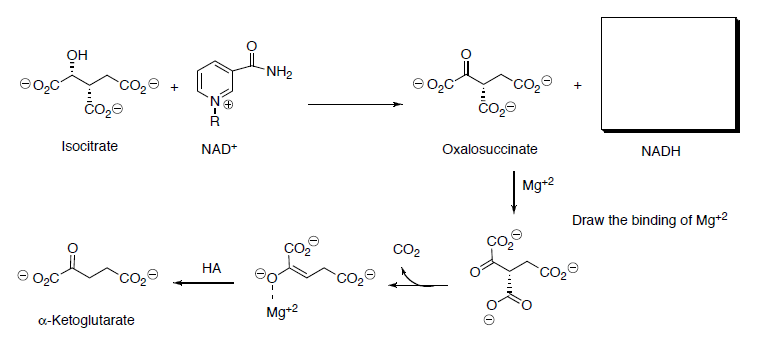
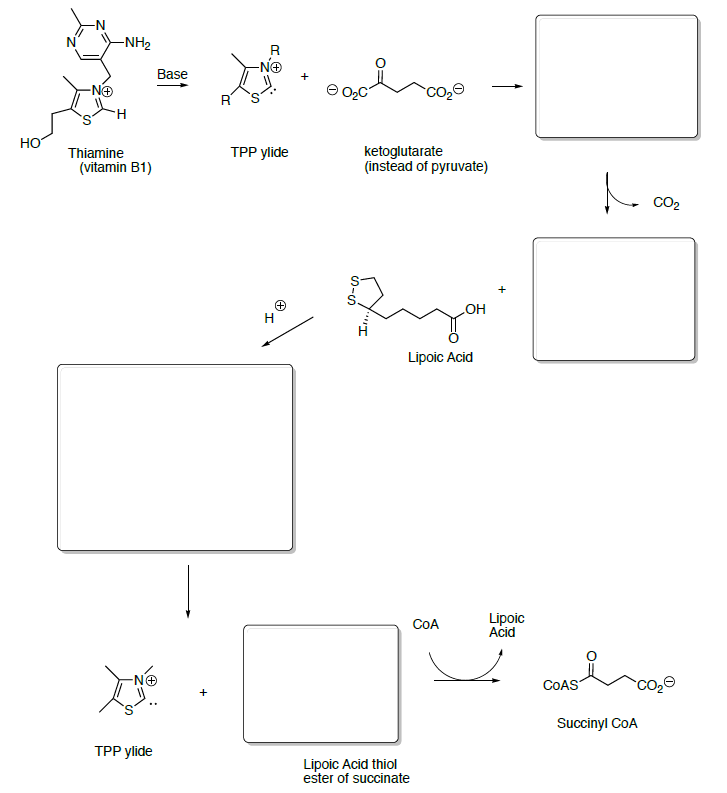
Ketoglutarate is transformed to succinyl CoA in a multistep process analogous to the transformation of pyruvate to acetyl CoA that we saw in the first step.
- Draw the transformation starting with the reaction with TPP ylide.
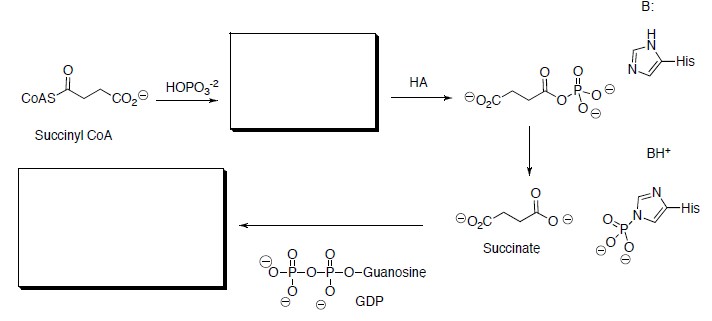
Succinyl CoA is hydrolyzed to succinate and is coupled with the phosphorylation of guanosine diphosphate (GDP) to give guanosine triphosphate (GTP).
- Draw the mechanism for this reaction.
Basic Metabolism: Oxidative Phosphorylation
Electron Transfer in Complex I
Complex I is located in the inner mitochondrial membrane in eukaryotes. The electrons from NADH (produced in the TCA cycle) begin to be shuttled through small steps to capture the energy.
This section will examine the mechanisms of electron transfer by the peripheral domain, proton transfer by the membrane domain and how their coupling can drive proton transport.
The net reaction of Complex I is the oxidation of NADH and the reduction of ubiquinone.
Net reaction:
\[\ce{NADH + H^+ + UQ \rightarrow NAD^+ + UQH2}\]
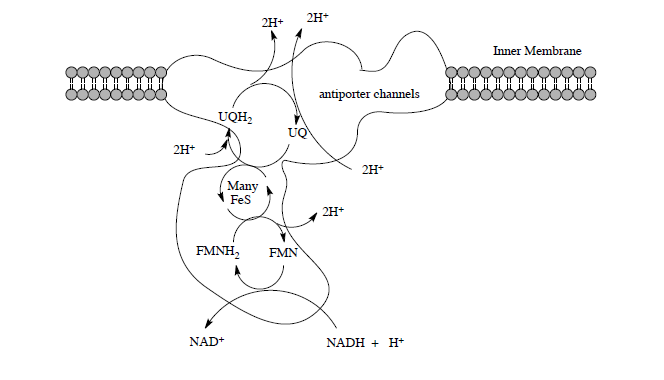
- How many protons are moved across the membrane for each cycle of Complex I?
- Is this active transport or passive diffusion?
- If this is active, what is fueling this transport?
- Is this with or against the concentration gradient? (i.e. antiporter or synporter?)
Complex II: Overview
Complex II (aka succinate dehydrogenase from the TCA cycle) oxidizes succinate (–O2CCH2CH2CO2–) to fumarate (trans-–O2CCH=CHCO2–).
Complex II also has a cascade of electron transfers. When succinate is converted to fumarate, the electrons are passed through a new cascade to eventually reduce UQ (just like Complex I!)
\[\ce{succinate \rightarrow fumarate + 2H+ + 2e-}\]
\[\ce{UQ + 2H+ + 2e- \rightarrow UQH2}\]
- Write the net reaction for the work of Complex II.
- The reaction catalyzed in Complex II has a very small ΔG°. Is it sufficient to power an antiporter channel?
Complex III: Overview
Complex III (sometimes called cytochrome bc1 complex) has two main substrates: cytochrome c and UQH2. The structure of this complex was determined by Johann Deisenhofer (Nobel Prize for a photosynthetic reaction center – we will see this soon).
This role of complex III is to transfer the electrons from UQH2 to cytochrome c.
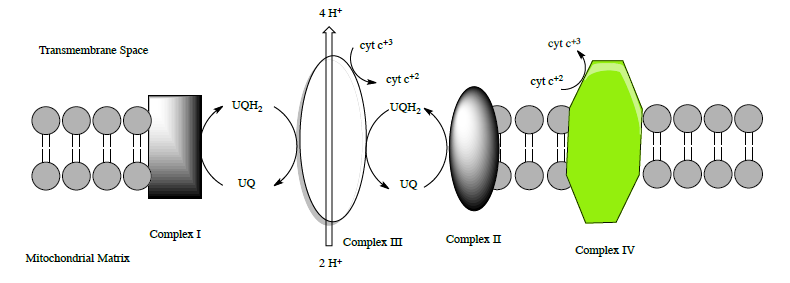
- Complete the equation for the redox reactions of complex III.
___ UQH2 + 1 UQ + 2 H+ + ___ cyt c+3 \(\ce{\rightarrow}\) ___ UQH2 + ___ UQ + 4 H+ + ___ cyt c+2
• There are two H+ coming from the mitochondrial matrix but _____ H+ are transported into the
inter-membrane space
Complex III to Complex IV: Cytochrome C as a mobile carrier
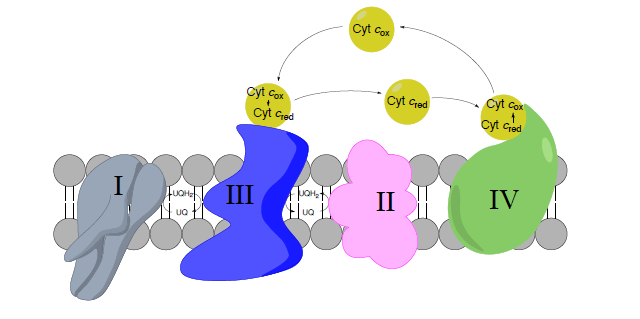
- Обведіть рухомі носії електронів на зображенні вище.
Огляд комплексу IV
Ще один комплекс, метою якого є переміщення електронів і протонів! Це великий крок, оскільки він є основним місцем для
утилізації диоксигену у всіх анаеробних організмах. Структура комплексу IV показана на малюнку зліва і праворуч на схемі, взятій з шляхів Кегга (з дозволу).
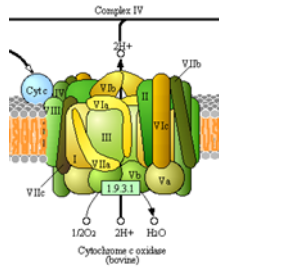
- Завершіть чисте рівняння для окисно-відновних реакцій комплексу IV.
___ цит с +2 + 1 О 2 + 8 Н +\(\ce{\rightarrow}\) ___ Н 2 О + 4 Н + ___ цит с +3
- Скільки протонів «закачується» в міжмембранний простір? _________
- Скільки електронів потрібно, щоб збалансувати це рівняння? _________
- Які початкові і кінцеві «рухливі» носії електронів?
Комплекс V: АТФ-синтаза
Нехтуючи Комплексом II, загальну реакцію мітохондріального ланцюга, на 2е — перенесену, можна записати як:
\[\ce{NADH + H+ + ½ O2 + 10 H+("in") \rightarrow NAD+ + H2O + 10 H+("out")} E° = +1.135V\]
Кожні два e — (від 1 молекули NADH) через ланцюг транспорту електронів призводить до чистого перенесення 10 протонів через мембрану:
- Комплекс I: ________ Н +
- Комплекс III: ________ Н +
- Комплекс IV: ________ Н +
Протони будуть дифундувати від області з високою концентрацією протонів до області нижчої концентрації протонів. Пітер Мітчелл отримав Нобелівську премію в 1978 році за свою пропозицію про те, що
електрохімічний градієнт концентрації протонів через мембрану може бути використаний для виготовлення АТФ. Протонний градієнт, створений ланцюгом транспорту електронів, забезпечує достатньо енергії для синтезу близько 2,5 молекул АТФ за допомогою процесу, званого хеміосмосом.
- Цей протонний потік приводиться в рух двома силами (заповнюють заготовки):
- Сила дифузії, викликана градієнтом концентрації. Всі частинки мають тенденцію рухатися від __________ концентрації до __________ концентрації.
- Електростатична сила, викликана градієнтом електричного потенціалу. Електричний градієнт є наслідком поділу заряду. Протони будуть залучені до ___________.
АТФ-синтаза є важливим ферментом, який використовує протонний градієнт привід синтезу (АТФ).
Електричний потенціал приводів двигуна
Ротор не заблокований у фіксованому положенні в центрі бішару і ділянки ротора перемикаються між порожнім і іонним зв'язаним станами. При русі синтезу АТФ з периплазми надходить іон і зв'язується на порожньому місці ротора.
Позитивний заряд статора (Arg 227) відіграє фундаментальну роль у функції двигуна F 0.
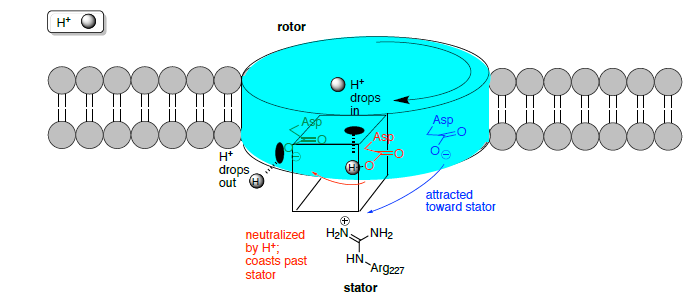
- Яка плата за порожні прив'язки сайтів:
- коли жоден іон не пов'язаний?
- коли зв'язується іон Na +/H +?
- Коли іон потрапляє на ділянку ротора з каналу статора, чистий заряд зменшується, таким чином [збільшуючи/зменшуючи] тяжіння до статора. Тепер ротор здатний переміщатися через гідрофобну частину статора, в той час як аргінін притягує наступний порожній ділянку ротора.
- Порожній ділянку (заряд = ________) електростатично притягується статором (заряд = ________) і направляється в наступну щілину.
- Місце ротора зайнято до тих пір, поки він не досягне статора з протилежного боку, де він стикається з позитивним зарядом статора, викликаючи дисоціацію іона. Чому? Розглянемо градієнти дифузії і заряди.
Електрична енергія Паливо Роторний АТФ Синтаза
- Заповніть пробіли на наступному резюме АТФ-синтази:
Під час синтезу АТФ градієнт ____________ підживлює вбудований в мембрану двигун F 0 для обертання центрального стебла. Це обертання викликає послідовні зміни зв'язування на периферійному домені F 1 так, що один каталітичний ділянку зв'язує ________ і фосфат, другий робить щільно пов'язаний АТФ, а третій крок ____________.
У анаеробно зростаючих бактерій, коли дихальні ферменти не активні, мотор F 1 може гідролізувати АТФ.
- В який бік повернеться насос в цих умовах?
- Що буде з двигуном F 0? А градієнт H +?
Джерела
Дімрот, Робота двигуна F0 синтази АТФ, Біохіміка та біофізика Acta (BBA) -Біоенергетика, 2000, 1458, 374-386.