3.2: Клітини нервової системи
- Page ID
- 90322

Психологи, які прагнуть зрозуміти людський розум, можуть вивчати нервову систему. Вивчення того, як функціонують клітини та органи організму, може допомогти нам зрозуміти біологічну основу психології людини. Нервова система складається з двох основних типів клітин: гліальних клітин (також відомих як глія) і нейронів. Традиційно вважається, що гліальні клітини відіграють підтримуючу роль нейронів, як фізично, так і метаболічно. Гліальні клітини забезпечують будівельні ліси, на яких побудована нервова система, допомагають нейронам тісно вибудовуватися один з одним, щоб забезпечити нейронний зв'язок, забезпечити ізоляцію нейронів, транспортувати поживні речовини та продукти життєдіяльності та опосередковувати імунні реакції. Протягом багатьох років дослідники вважали, що гліальних клітин набагато більше, ніж нейронів; однак, більш пізня робота лабораторії Сюзанни Геркулано-Хузель поставила це давнє припущення під сумнів і надала важливі докази того, що може бути майже 1:1 співвідношення клітин глії до нейронів. Це важливо, оскільки це говорить про те, що людський мозок більше схожий на інші мізки приматів, ніж вважалося раніше (Azevedo et al, 2009; Hercaulano-Houzel, 2012; Herculano-Houzel, 2009). Нейрони, з іншого боку, служать взаємопов'язаними інформаційними процесорами, які є важливими для всіх завдань нервової системи. У цьому розділі коротко описується будова і функції нейронів.
Структура нейрона
Нейрони є центральними будівельними блоками нервової системи, 100 мільярдів сильних при народженні. Як і всі клітини, нейрони складаються з декількох різних частин, кожна виконує спеціалізовану функцію (рис. 3.8). Зовнішня поверхня нейрона складається з напівпроникної мембрани. Ця мембрана дозволяє меншим молекулам і молекулам без електричного заряду проходити через неї, зупиняючи при цьому більші або сильно заряджені молекули.
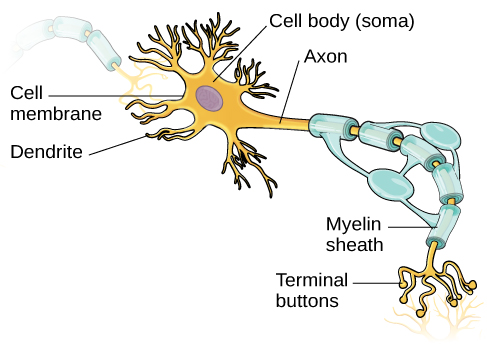
Ядро нейрона розташовується в сомі, або тілі клітини. Сома має розгалужені розширення, відомі як дендрити. Нейрон є невеликим інформаційним процесором, а дендрити служать вхідними ділянками, куди надходять сигнали від інших нейронів. Ці сигнали передаються електрично через сому і вниз великим розширенням від соми, відомої як аксон, який закінчується на декількох термінальних кнопках. Кінцеві кнопки містять синаптичні везикули, в яких розміщені нейромедіатори, хімічні посланники нервової системи.
Аксони варіюються в довжину від частки дюйма до декількох футів. У деяких аксонів гліальні клітини утворюють жирну речовину, відому як мієлінова оболонка, яка покриває аксон і діє як ізолятор, збільшуючи швидкість, з якою рухається сигнал. Мієлінова оболонка не є безперервною, і є невеликі проміжки, які виникають вниз по довжині аксона. Ці прогалини в мієлінової оболонці відомі як Вузли Ранв'є. Мієлінова оболонка має вирішальне значення для нормальної роботи нейронів у нервовій системі: втрата ізоляції, яку вона забезпечує, може згубно позначитися на нормальній роботі. Щоб зрозуміти, як це працює, розглянемо приклад. ФКУ, генетичне захворювання, про яке йшлося раніше, викликає зниження мієліну та аномалії білої речовини коркових і підкіркових структур. Розлад пов'язаний з різними проблемами, включаючи важкий когнітивний дефіцит, перебільшені рефлекси та судоми (Anderson & Leuzzi, 2010; Huttenlocher, 2000). Інший розлад, розсіяний склероз (РС), аутоімунний розлад, передбачає масштабну втрату мієлінової оболонки на аксони по всій нервовій системі. Внаслідок цього перешкоди в електричному сигналі перешкоджають швидкій передачі інформації нейронами і можуть призвести до ряду симптомів, таких як запаморочення, втома, втрата рухового контролю та сексуальна дисфункція. Хоча деякі методи лікування можуть допомогти змінити перебіг захворювання та керувати певними симптомами, в даний час не існує відомих ліків від розсіяного склерозу.
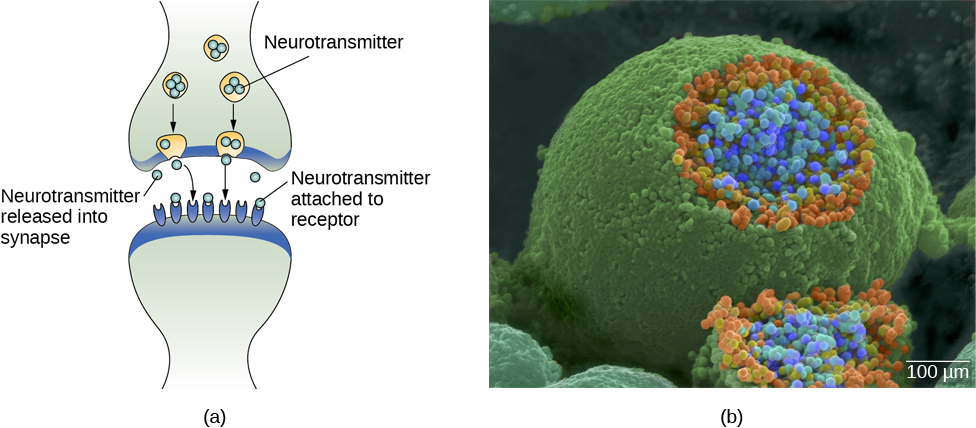
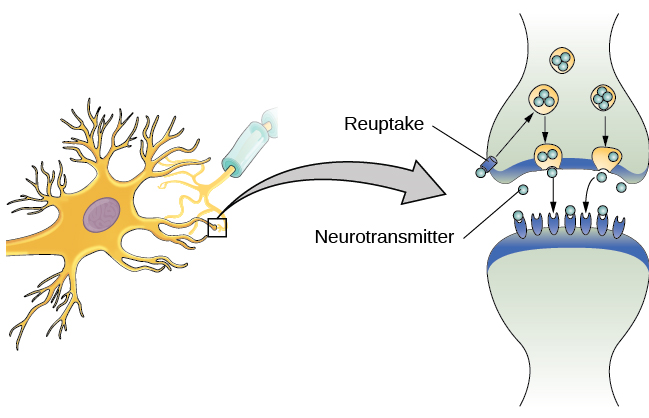
У здорових осіб нейрональний сигнал швидко рухається вниз по аксону до кінцевих кнопок, де синаптичні везикули випускають нейромедіатори в синаптичну щілину (рис. 3.9). Синаптична щілина - це дуже маленький простір між двома нейронами і є важливим місцем, де відбувається зв'язок між нейронами. Як тільки нейромедіатори вивільняються в синаптичну щілину, вони подорожують по ній і зв'язуються з відповідними рецепторами на дендриті сусіднього нейрона. Рецептори, білки на поверхні клітини, де прикріплюються нейромедіатори, змінюються за формою, з різними формами «відповідають» різним нейромедіаторам.
Як нейромедіатор «знає», з яким рецептором зв'язуватися? Нейромедіатор і рецептор мають те, що називається блокуванням та ключовим зв'язком - специфічні нейромедіатори підходять конкретним рецепторам, подібним до того, як ключ підходить до блокування. Нейромедіатор зв'язується з будь-яким рецептором, який він підходить.
Нейрональна комунікація
Тепер, коли ми дізналися про основні структури нейрона та роль, яку ці структури відіграють у нейронному спілкуванні, давайте детальніше розглянемо сам сигнал - як він рухається через нейрон, а потім переходить до наступного нейрона, де процес повторюється.
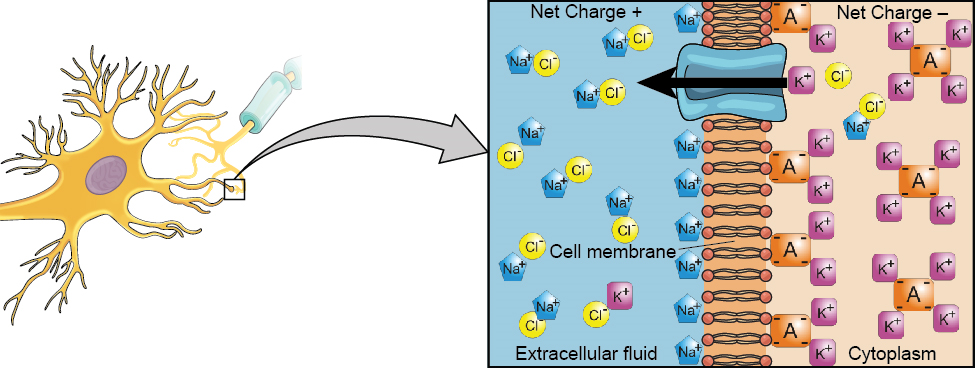
Починаємо з нейронної мембрани. Нейрон існує в рідкому середовищі - він оточений позаклітинною рідиною і містить внутрішньоклітинну рідину (тобто цитоплазму). Нейронна мембрана тримає ці дві рідини окремо - вирішальну роль, оскільки електричний сигнал, який проходить через нейрон, залежить від електрично різних внутрішньо-та позаклітинних рідин. Ця різниця заряду по мембрані, звана мембранним потенціалом, забезпечує енергію для сигналу.
Електричний заряд рідин викликається зарядженими молекулами (іонами), розчиненими в рідині. Напівпроникний характер мембрани нейронів дещо обмежує рух цих заряджених молекул, і, як наслідок, частина заряджених частинок має тенденцію ставати більш концентрованими або всередині, або зовні клітини.
Між сигналами потенціал мембрани нейрона утримується в стані готовності, званому потенціалом спокою. Як гумова стрічка, розтягнута і чекає, щоб весна вступила в дію, іони шикуються по обидва боки клітинної мембрани, готові кидатися через мембрану, коли нейрон стає активним і мембрана відкриває свої ворота (тобто натрієво-калієвий насос, який дозволяє рух іонів через мембрану). Іони в районах з високою концентрацією готові переміщатися в області низької концентрації, а позитивні іони готові переміщатися в області з негативним зарядом.
У стані спокою натрій (Na +) знаходиться в більш високих концентраціях поза клітиною, тому він буде схильний рухатися в клітину. Калій (K +), з іншого боку, більш концентрований всередині клітини і буде мати тенденцію рухатися з клітини (рис. 3.10). Крім того, внутрішня частина клітини трохи негативно заряджена в порівнянні з зовнішньою. Це забезпечує додаткове зусилля на натрій, змушуючи його переміщатися в клітину.
З цього стану потенціалу спокою нейрон отримує сигнал і його стан різко змінюється (рис. 3.11). Коли нейрон отримує сигнали на дендритах - завдяки нейромедіаторам від сусіднього нейрона, що зв'язується з його рецепторами - дрібні пори або ворота, відкриті на мембрані нейронів, дозволяючи іонам Na +, що рухаються як зарядом, так і різницею концентрацій, рухатися в клітину. При такому припливі позитивних іонів внутрішній заряд клітини стає більш позитивним. Якщо цей заряд досягає певного рівня, званого порогом збудження, нейрон стає активним і починається потенціал дії.
Багато додаткових пір відкриваються, викликаючи масовий приплив іонів Na + і величезний позитивний сплеск мембранного потенціалу, пікового потенціалу дії. На піку шипа натрієві ворота закриваються, а калієві ворота відкриваються. Оскільки позитивно заряджені іони калію йдуть, клітина швидко починає реполяризацію. Спочатку він гіперполяризується, стаючи трохи більш негативним, ніж потенціал спокою, а потім вирівнюється, повертаючись до потенціалу спокою.
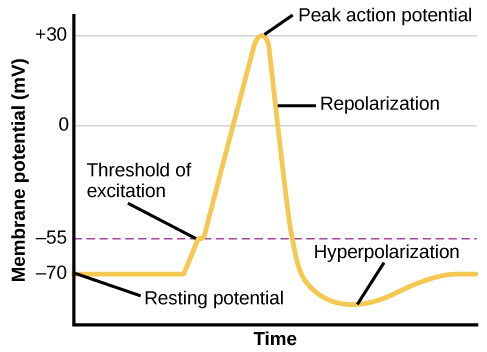
Цей позитивний сплеск становить потенціал дії: електричний сигнал, який зазвичай рухається від тіла клітини вниз по аксону до клем аксона. Електричний сигнал рухається вниз по аксону з імпульсами, що стрибають чехарда між вузлами Ранв'є. Вузли Ранв'є є природними прогалинами в мієлінової оболонці. У кожній точці частина іонів натрію, що надходять в клітину, дифузіруется в наступну секцію аксона, піднімаючи заряд повз поріг збудження і запускаючи новий приплив іонів натрію. Потенціал дії таким чином рухається вниз по аксону, поки не дійде до кінцевих кнопок.
Потенціал дії - це явище «все або ні». Говорячи простою мовою, це означає, що вхідний сигнал від іншого нейрона або достатній, або недостатній для досягнення порога збудження. Між ними немає, і немає вимкнення потенціалу дії після його запуску. Подумайте про це, як надсилання електронного листа або текстового повідомлення. Ви можете думати про відправку все, що хочете, але повідомлення не відправляється, поки ви не натиснете кнопку відправити. Крім того, як тільки ви надішлете повідомлення, його немає зупинки.
Оскільки це все або ні, потенціал дії відтворюється або поширюється на повну силу в кожній точці вздовж аксона. Подібно до запаленого запобіжника петарди, він не згасає, коли рухається вниз по аксону. Саме ця властивість «все або ні» пояснює той факт, що ваш мозок сприймає травму віддаленої частини тіла, як ваш палець ноги так само болісно, як один до вашого носа.
Як зазначалося раніше, коли потенціал дії надходить на кінцеву кнопку, синаптичні бульбашки випускають свої нейромедіатори в синаптичну щілину. Нейромедіатори подорожують по синапсу і зв'язуються з рецепторами на дендритах сусіднього нейрона, і процес повторюється в новому нейроні (припускаючи, що сигнал досить сильний, щоб викликати потенціал дії). Після доставки сигналу надлишкові нейромедіатори в синаптичній щілині відходять, розбиваються на неактивні фрагменти або реабсорбуються в процесі, відомому як зворотне захоплення. Зворотне захоплення включає в себе нейромедіатор, який перекачується назад в нейрон, який випустив його, щоб очистити синапс (рис. 3.12). Очищення синапсу служить як для забезпечення чіткого стану «увімкнено» та «вимкнено» між сигналами, так і для регулювання виробництва нейромедіатора (повні синаптичні везикули дають сигнали про те, що додаткові нейромедіатори не потрібно виробляти).
Нейромедіатори та наркотики
Існує кілька різних типів нейромедіаторів, що виділяються різними нейронами, і ми можемо говорити в широких рисах про види функцій, пов'язаних з різними нейромедіаторами (табл. 3.1). Багато що з того, що психологи знають про функції нейромедіаторів, походить від досліджень впливу наркотиків при психологічних розладах. Психологи, які приймають біологічну перспективу і зосереджуються на фізіологічних причині поведінки, стверджують, що психологічні розлади, такі як депресія та шизофренія, пов'язані з дисбалансами в одній або декількох нейромедіаторних системах. У цій перспективі психотропні препарати можуть допомогти покращити симптоми, пов'язані з цими розладами. Психотропні препарати - це препарати, які лікують психіатричні симптоми шляхом відновлення балансу нейромедіаторів.
| Нейромедіатор | Причетний до | Потенційний вплив на поведінку |
|---|---|---|
| ацетилхолін | Дія м'язів, пам'ять | Підвищене збудження, посилене пізнання |
| Бета-ендорфін | Біль, задоволення | Зниження тривожності, зниження напруги |
| Дофамін | Настрій, сон, навчання | Підвищене задоволення, пригнічений апетит |
| Гамма-аміномасляна кислота (ГАМК) | Функція мозку, сон | Зниження тривожності, зниження напруги |
| глутамат | Пам'ять, навчання | Збільшення навчання, покращена пам'ять |
| норадреналін | Серце, кишечник, настороженість | Підвищене збудження, пригнічений апетит |
| серотонін | Настрій, сон | Модульований настрій, пригнічений апетит |
Психоактивні препарати можуть виступати агоністами або антагоністами для даної системи нейромедіаторів. Агоністи - це хімічні речовини, які імітують нейромедіатор на місці рецептора. Антагоніст, з іншого боку, блокує або перешкоджає нормальній діяльності нейромедіатора на рецепторі. Агоністи та антагоністи являють собою препарати, які призначаються для корекції специфічних дисбалансів нейромедіаторів, що лежать в основі стану людини. Наприклад, хвороба Паркінсона, прогресуюче розлад нервової системи, пов'язана з низьким рівнем дофаміну. Тому загальна стратегія лікування хвороби Паркінсона передбачає використання агоністів дофаміну, які імітують ефекти дофаміну шляхом зв'язування з дофаміновими рецепторами.
Певні симптоми шизофренії пов'язані з гіперактивною нейротрансмісією дофаміну. Антипсихотики, що використовуються для лікування цих симптомів, є антагоністами дофаміну - вони блокують ефекти дофаміну, зв'язуючи його рецептори, не активуючи їх. Таким чином, вони перешкоджають дофаміну, що виділяється одним нейроном, сигналізувати інформацію сусіднім нейронам.
На відміну від агоністів та антагоністів, які обидва діють шляхом зв'язування з рецепторними сайтами, інгібітори зворотного захоплення запобігають транспортуванню невикористаних нейромедіаторів назад до нейрона. Це дозволяє нейромедіаторам залишатися активними в синаптичній щілині протягом більшої тривалості, підвищуючи їх ефективність. Депресія, яка послідовно пов'язана зі зниженим рівнем серотоніну, зазвичай лікується селективними інгібіторами зворотного захоплення серотоніну (СИОЗС). Запобігаючи зворотному захопленню, СИОЗС підсилюють дію серотоніну, даючи йому більше часу для взаємодії з рецепторами серотоніну на дендритах. Поширені СИОЗС на ринку сьогодні включають Прозак, Паксил і Золофт. Препарат ЛСД структурно дуже схожий на серотонін, і він впливає на ті ж нейрони і рецептори, що і серотонін. Психотропні препарати не є миттєвими рішеннями для людей, які страждають психологічними розладами. Часто людина повинна приймати препарат протягом декількох тижнів, перш ніж побачити поліпшення, і багато психоактивних препаратів мають значні негативні побічні ефекти. Крім того, люди різко варіюються в тому, як вони реагують на наркотики. Щоб покращити шанси на успіх, не рідкість люди, які отримують фармакотерапію, також проходять психологічну та/або поведінкову терапію. Деякі дослідження показують, що поєднання медикаментозної терапії з іншими формами терапії має тенденцію бути більш ефективним, ніж будь-яке одне лише лікування (для одного з таких прикладів див. March et al., 2007).