8: Відповідь на фенотипічний відбір
- Page ID
- 7754
Еволюція шляхом природного відбору вимагає:
- Варіація фенотипу
- Це виживання та розмноження є невипадковими щодо цієї фенотипової варіації.
- Що ця варіація є спадковою.
Пункти 1 і 2 інкапсулюють нашу ідею природного відбору, але еволюція шляхом природного відбору відбудеться лише в тому випадку, якщо буде виконана і 3-я умова. Це спадкова природа варіацій, що пари змінюються протягом покоління завдяки природному відбору, щоб змінюватися між поколіннями (еволюційні зміни).
Почнемо з того, що подумаємо про зміну всередині покоління через спрямований відбір, де відбір діє на зміну середнього фенотипу в поколінні. Наприклад, зменшення середньої висоти протягом покоління через більш високі організми, які мають менший шанс вижити до розмноження, ніж коротші організми. Зокрема, ми позначимо наш середній фенотип при розмноженні\(\mu_S\), тобто після відбору діяв, і наш середній фенотип перед відбором діє\(\mu_{BS}\). Цю другу величину може бути важко виміряти, оскільки очевидно, що відбір діє протягом усього життєвого циклу, тому було б легше думати про це як про середній фенотип, якщо відбір не діяв. Таким чином, зміна середнього фенотипу всередині покоління\(\mu_{S} - \mu_{BS}= S\), ми будемо називати\(S\) вибір диференціал.
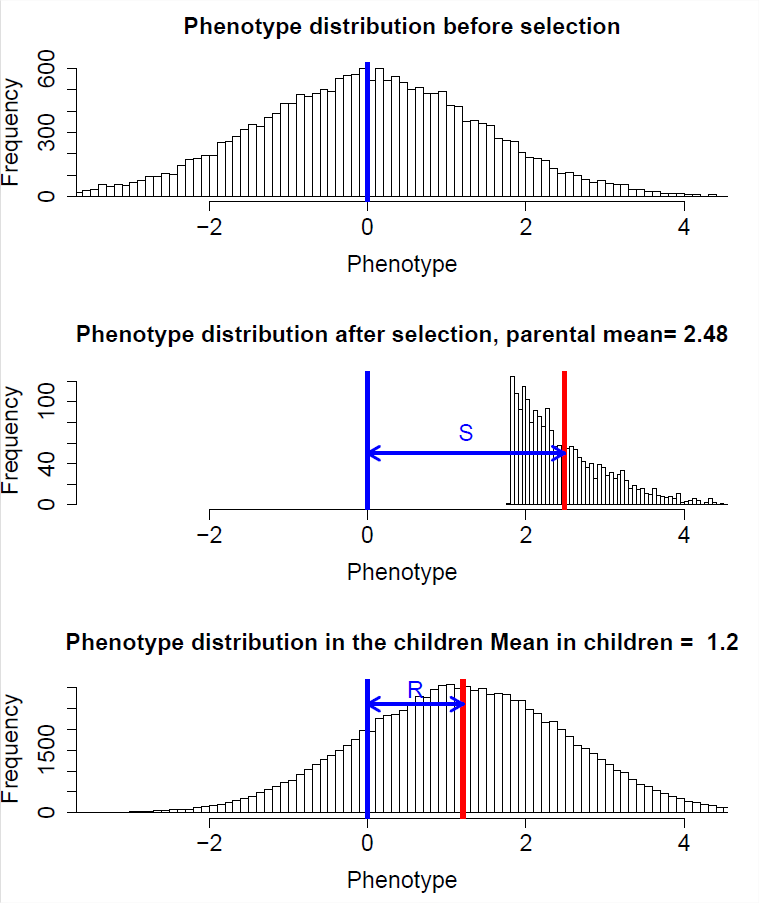
Ми зацікавлені в прогнозуванні розподілу фенотипів у наступному поколінні. Зокрема, нас цікавить середній фенотип у наступному поколінні, щоб зрозуміти, як спрямований відбір сприяв еволюційним змінам. Ми позначимо середній фенотип у потомстві, тобто середній фенотип у наступному поколінні перед відбором діє, як\(\mu_{NG}\). Зміни між поколіннями ми назвемо відповіддю на вибір\(R\) і поставимо це рівним\(\mu_{NG}- \mu_{BS}\).
Середній фенотип у наступному поколінні
\[\mu_{NG} = \E \left( \mathbb{E}(X_{kid} | X_{mum},X_{dad}) \right)\]
де внутрішнє очікування дає нам очікуваний фенотип дитини, що дається батькам, а зовнішнє очікування перевищує можливі пари батьків, утворених випадковим спаровуванням особин, які виживають для розмноження. Ми можемо використовувати Equation\ ref {predict_kid}, щоб отримати вираз для цього очікування:
\[\mu_{NG} = \mu_{BS} + \beta_{mid,kid} ( \mathbb{E}(X_{mid}) - \mu_{BS})\]
Отже, для отримання\(\mu_{NG}\) нам потрібно\(\mathbb{E}(X_{mid})\) обчислити очікуваний фенотип середньої точки пар особин, які виживають до розмноження. Це якраз очікуваний фенотип у особин, які вижили до розмноження (\(\mu_{S}\)), тому
\[\mu_{NG} = \mu_{BS} + h^2 (\mu_S - \mu_{BS})\]
Таким чином, ми можемо написати нашу відповідь на вибір як
\[R = \mu_{NG} -\mu_{BS} = h^2 (\mu_S - \mu_{BS}) = h^2 S \label{breeders_eqn}\]
Отже, наша реакція на вибір пропорційна нашому диференціалу вибору, а константа пропорційності - це успадкованість вузького смислу. Це рівняння іноді називають рівнянням селекціонера. Це твердження, що еволюційна зміна між поколіннями (\(R\)) пропорційна зміні, спричиненій спрямованим відбором всередині покоління (\(S\)), і що сила цього зв'язку визначається вузьким смислом спадковості ( \(h^2\)).
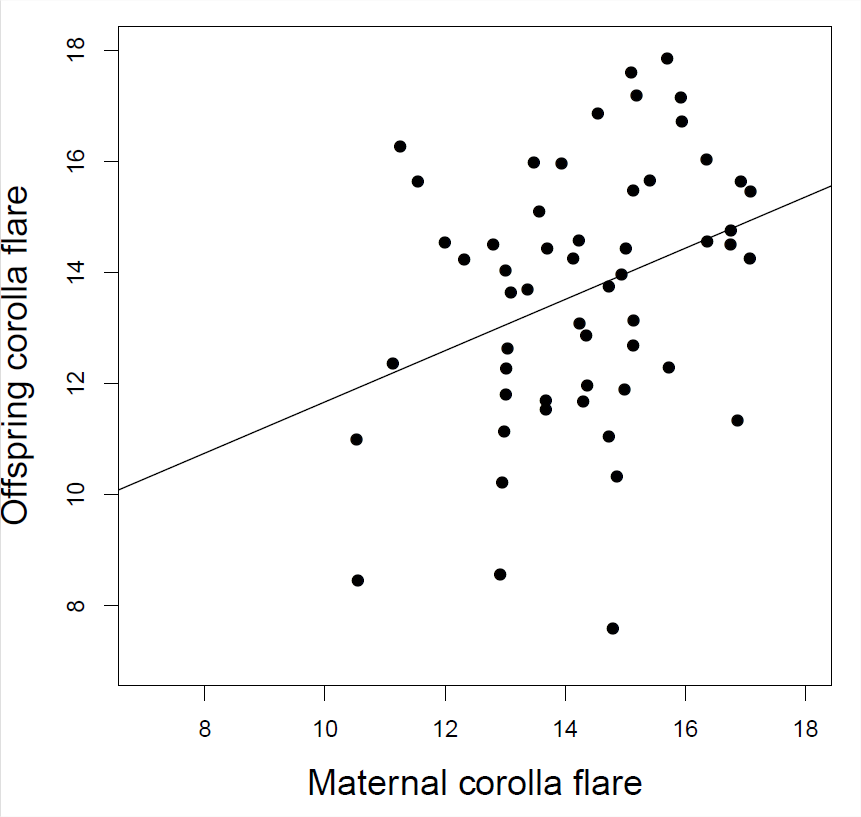
Гален досліджував селекцію за формою квітки у Polemonium viscosum. Вона виявила, що рослини з більшим відблиском віночка мали більше відвідувань джмелів, що призвело до більш високого набору насіння та\(17\%\) збільшення спалаху віночка у рослин, що сприяють наступному поколінню. Виходячи з даних у заголовку Figure\ ref {fig:Galen_Corolla} яка очікувана відповідь у наступному поколінні?

Якщо ми знаємо\(R\) і\(S\) можемо оцінити\(h^2\). Спадковість, оцінена так, називається «реалізованою спадщиною». Оцінки «реалізованої спадщини» можуть бути легко отримані в експериментах з штучним відбором:
З експерименту, показаного на малюнку\ ref {Fig:illinois_LTS_Breeders_EQ}, середній вміст кукурудзяної олії в 1897 році був серед\(24\) особин\(4.78\), обраних для розведення для наступного покоління, середнє значення було\(5.2\). Нащадки цих особин мали середнє ядро олійності\(5.1\). У чому вузькому сенсі реалізована спадковість?
Щоб зрозуміти генетичну основу відповіді на селекцію, зверніть увагу на рис.\ ref {Fig:response_num_alleles}. Налаштування така ж, як і в наших попередніх моделювальних малюнках.

Особи, які відібрані для формування нашого наступного покоління, несуть більше алелів, які збільшують фенотип у поточному діапазоні середовищ, які зараз переживає населення. Середній індивід перед відбором проводив 100 з цих алелів «вгору», тоді як середній індивідуальний вижив відбір несе 108 алелів «вгору».
Оскільки особи вірно передають свої алелі наступному поколінню, середня дитина обраних батьків несе\(108\) алелі. Зауважте, що дисперсія мало змінилася, діти мають багато варіацій у своєму генотипі, так що селекція може легко керувати еволюцією в майбутніх поколіннях. Середня частота алеля «вгору» змінилася від\(50\%\) до\(54\%\). Прибутки завдяки відбору будуть стабільно успадковуватися майбутнім поколінням і можуть бути посилені на покоління за поколінням, якщо тиск на відбір залишатиметься постійним.
Довгострокова реакція на відбір
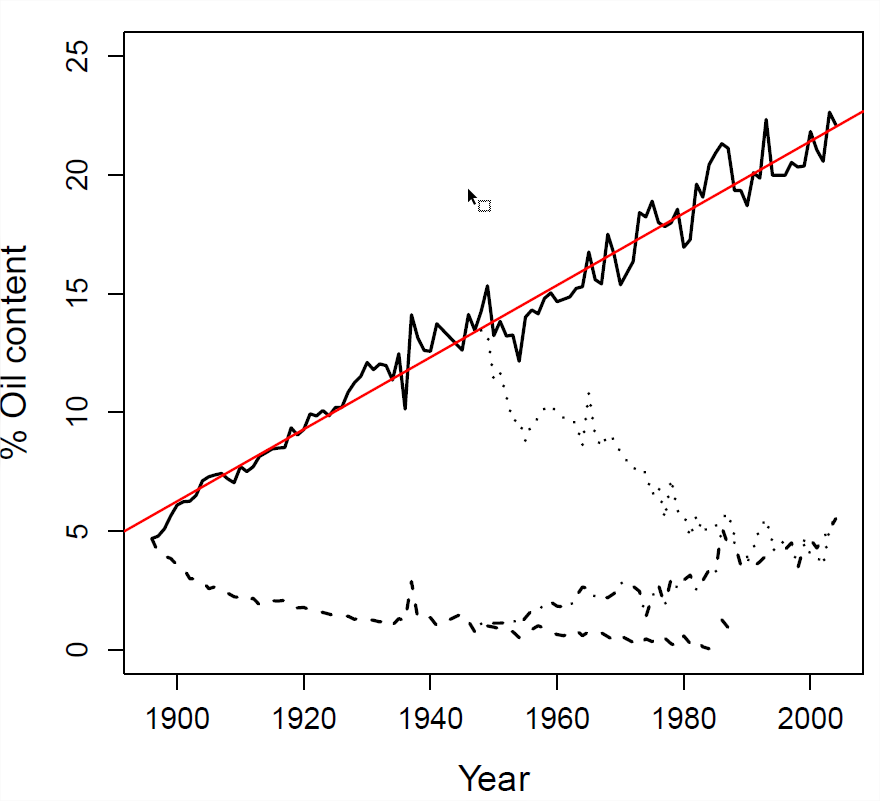
Лінійна підгонка до експерименту показана червоною лінією. Дані доступні https://www.ideals.illinois.edu/handle/2142/3525, https://github.com/cooplab/popgen-no...orn/corn_LTS.R
Якщо наш селекційний тиск підтримується протягом багатьох поколінь, ми можемо використовувати рівняння нашого селекціонера для прогнозування реакції. Якщо ми готові припустити, що наша спадковість не змінюється, і ми підтримуємо постійний диференціал відбору (\(S\)), то після\(n\) поколінь наше середнє значення фенотипу зміститься
\[n h^2 S\]
тобто наше населення буде підтримувати лінійну реакцію на відбір.
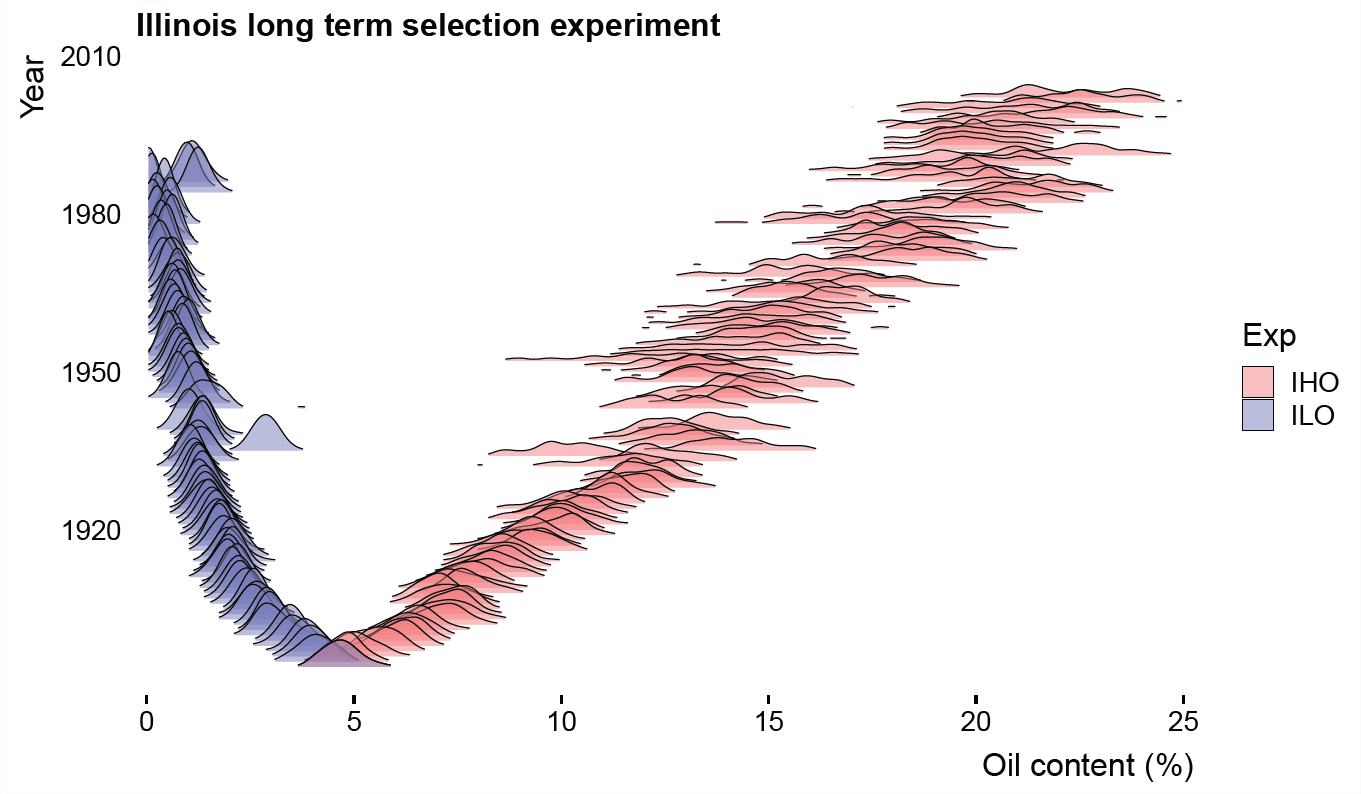
Тому довгостроковий послідовний відбір може призвести до вражаючих еволюційних змін. Одним із прикладів цього є польовий експеримент в штаті Іллінойс, де селекціонери рослин систематично вибирали вищий і менший вміст олії в кукурудзі (див. Наш попередній малюнок\ ref {Fig:illinois_LTS_Breeders_EQ} для одного покоління вгору селекції). Понад століття вони беруть насіння у рослин в крайніх межах поширення і використовують їх для формування наступного покоління. Вони досягли вражаючих довгострокових відповідей, виштовхуючи розподіл населення далеко за межі їх початкового діапазону (рис.\ ref {Fig:illinois_LTS_dists}. Наприклад, лінія підбору нафти пішла від середнього вмісту нафти\(4.7\%\) в 1896\(22.1\%\) році до 2004 року. Вони встановили дві вторинні популяції, де диференціал відбору був скасований. У популяції вищого відбору вони підтримували вражаюче лінійне збільшення вмісту олії, показане червоною лінією на малюнку\ ref {Fig:illinois_LTS_means}, але хоча реакція спочатку лінійна в нижній лінії, але вони швидко досягають дуже низького вмісту олії (обмежена фізичною межею при 0% вмісту олії).
Популяція благородних оленів опинилася в пастці на Джерсі (острів біля Англії) протягом останнього міжльодовикового періоду. З викопного запису ми бачимо, що популяція швидко адаптувалася до своїх нових умов, можливо, завдяки відбору на більш короткий час розмноження за відсутності хижацтва. Протягом 6000 років вони еволюціонували від орієнтовної середньої ваги населення 200 кг до орієнтовної середньої ваги 36 кг (зменшення в 6 разів)! Ви підрахували, що час генерації благородного оленя становить 5 років, і з поточного дня популяції, що вузький сенс спадковості фенотипу становить 0,5.
- Оцініть середню зміну на покоління в середній масі тіла.
- Оцініть зміну середньої маси тіла, спричинену відбором протягом покоління. Викладіть свої припущення.
- Припускаючи, що ми маємо лише скам'янілості від населення-засновника та населення після 6000 років, чи варто припускати, що розрахунки точно відображають те, що насправді сталося в нашому населенні?
Санта-Роза {\ це Маммут exilis}. \ href {https://en.Wikipedia.org/wiki/Pygmy_...s_skeletal.png, CC BY 3.0.
У диких популяціях тиск на відбір, ймовірно, рідко підтримується для великої кількості поколінь. Наприклад, Гранти вимірювали фенотипічний відбір у В'юрків Дарвіна протягом декількох десятиліть на острові Дафна Майор. Вони бачили, що тиск відбору в Середньому в'юрку (Geospiza fortis) змінювався кілька разів протягом багатьох років (Рис.\ ref {рис:Darwins_Finches_unpred}).
Закономірності довгострокових фенотипічних змін у дикій природі
Дивлячись на різноманітність рослин і тварин, ми бачимо величезні зміни в розмірах і формі, чи можуть сильні сторони відбору, які ми можемо спостерігати за короткі періоди часу, можливо, пояснити ці зміни?
Для порівняння фенотипічних змін протягом різних часових періодів нам потрібна певна міра швидкості фенотипічної зміни. запропонована швидкість зміни від\(X_1\) до\(X_2\) часового інтервалу\(\Delta t\), вимірювана мільйонами років, кількісно оцінюється як
\[\frac{\log \left(\frac{X_2}{X_1} \right) }{\Delta t} = \frac{\log \left(X_2 \right) -\log \left(X_1 \right) }{\Delta t}\]
висловлюючи це журнал співвідношення, ми дивимося на пропорційну зміну складки, що має сенс, оскільки еволюційна зміна довжини 1 см є більш вражаючою, якщо ви миша, ніж слон. Помістивши це на\(\log\) -шкалу, ми дивимося на відносну зміну. називається одиниці цієї міри `Дарвіна ', з однією зміною Дарвіна, що відповідає зміні\(e\approx 2.71\) згину за мільйон років, дві зміни Дарвіна, що відповідають \(e^2\approx 7.34\)складка зміна через мільйон років і так далі.
Обчисліть швидкість зміни розміру тіла в Джерсі благородного оленя з питання\ ref {question:reddeer} в Darwins. Зробіть те ж саме для загальної зміни вмісту кукурудзяної олії в рядках вгору на малюнку\ ref {рис:illinois_LTS_means}.
досліджено абсолютну швидкість фенотипічних змін даних польових досліджень та запису викопних копалин, набір даних значно розширений на. На малюнку\ ref {fig:uyeda_gingerich} кожна точка є спостереженням еволюції фенотипу. Вісь x показує часовий проміжок у роках, протягом якого спостерігалися еволюційні зміни, вісь x - побудована за\(\log_{10}\) шкалою. Вісь y показує абсолютну швидкість фенотипічних змін, виміряну в Дарвіні, знову ж таки за\(\log_{10}\) шкалою.
За короткі часові шкали ми спостерігаємо неймовірно швидку еволюцію, зверніть увагу на високі показники зліва від малюнка\ ref {fig:uyeda_gingerich}. Наприклад, перша чорна крапка зліва - це випадок еволюції протягом десятиліть у собачих щиків. Вторгнення зеленого краба (Carcinus maenas) призвело до еволюції більш міцних снарядів в атлантичному собачому колесі (Nucella lapillus) у відповідь на хижацтво уздовж північноамериканського узбережжя. Товщина губи оболонки собак у Сент-Ендрюсі, населення Нью-Брансвік змінилася з 0,94 мм до 1,44 мм всього за 25 років. Це 50% збільшення, і ставка 17060 Дарвінів.
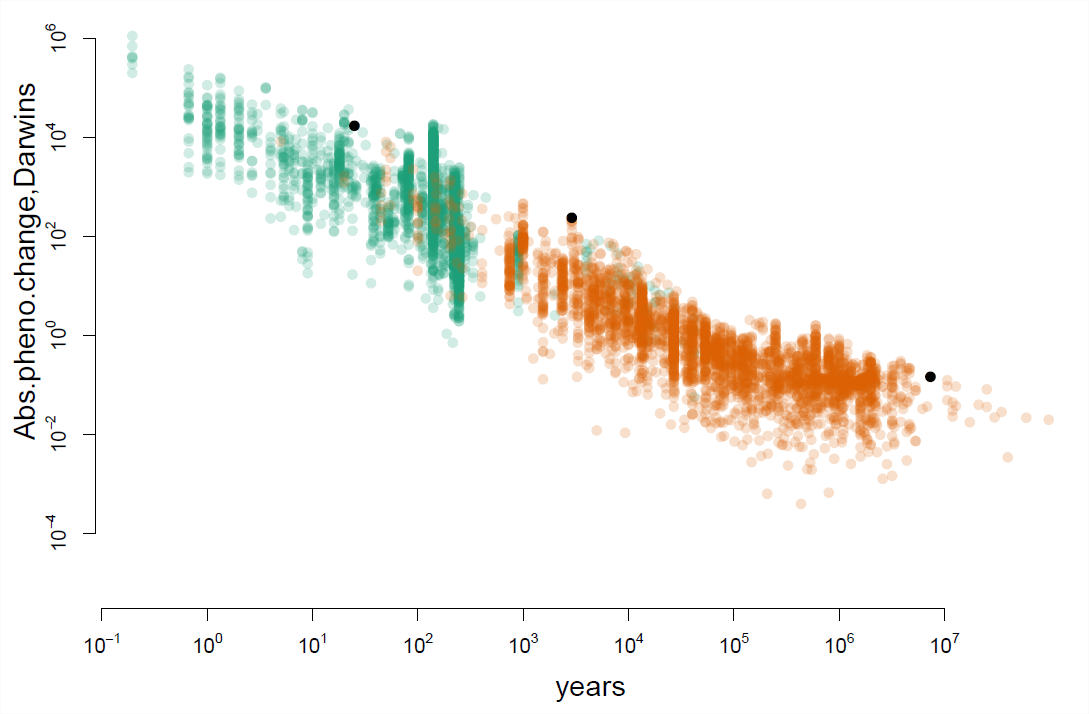
Однак, коли ми спостерігаємо фенотипічну еволюцію протягом більш тривалих періодів часу, це, як правило, набагато повільніше. Наприклад, крайня права чорна точка на малюнку\ ref {fig:uyeda_gingerich} показує фенотипічну еволюцію вздовж лінії, що веде до трицератопсу. Трицератопс вимірюється у вражаючій довжині 25,9 - 29,5 футів. Вони еволюціонували від близького родича протоцератопса, який був трохи більшим за вівцю на\(\sim\) 5,9 футів приблизно за 7,5 мільйона років. Однак це лише фенотипічна зміна\(0.143\) Дарвіна, це лише приблизно чотириразова зміна за мільйони років. Ці темпи змін у динозаврів нічого не мають на наших собачих вагах або багатьох інших прикладах еволюції на коротких часових масштабах. Таким чином, еволюційні зміни, які ми можемо спостерігати за короткими часовими шкалами, легко пояснюють довгострокові зміни кількісних фенотипів.
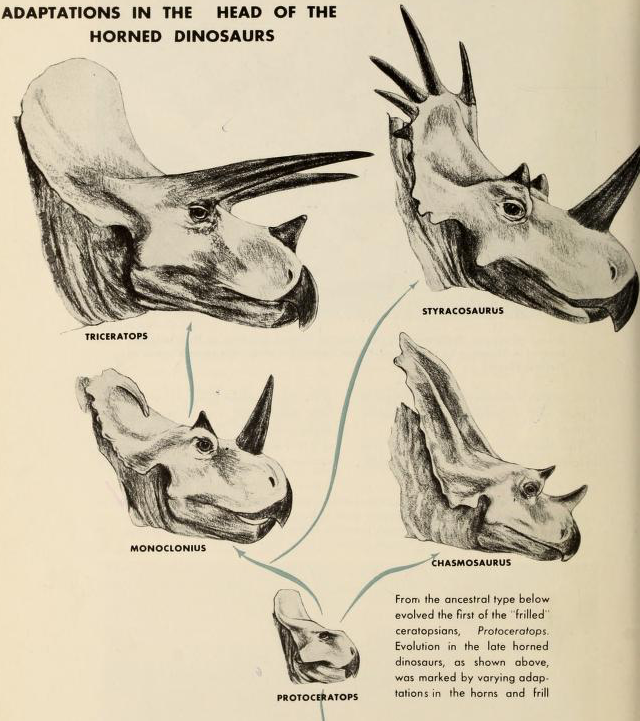
для цікавого оновленого уявлення про філогенію {\ it Coronosauria}. Дивіться ці
\ href {https://www.geol.umd.edu/~tholtz/G10...04margino.html від Holtz для оновленої\ & повної філогенії. \ IANC {Книга динозаврів: правлячі рептилії та їх родичі. (1951) Кольбер, Е.Х. https://www.biodiversitylibrary.org/.../1up}{American Музей природничої бібліотеки
Фітнес та рівняння заводчика.
Отже, спрямована еволюція відбувається, коли селекція призводить до зміни середнього фенотипу в межах покоління. Але як саме це пов'язано з вимогою природного відбору, що організми змінюються у своїй придатності? Деякі різні способи формулювання рівняння селекціонера дають нам уявлення про умови спрямованого відбору та відношення до фітнес-ландшафтів.
Спрямований відбір як коваріація між фітнесом і фенотипом.
Щоб більш ретельно подумати про цю зміну протягом покоління, давайте подумаємо про просту фітнес-модель, де наш фенотип впливає на життєздатність наших організмів (тобто ймовірність того, що вони виживуть для розмноження). Імовірність того, що індивід має фенотип\(X\) перед відбором\(p(X=x)\), є, так що середній фенотип перед відбором
\[\mu_{BS} = \E[X] = \int_{-\infty}^{\infty} x p(x) dx\]
Імовірність того, що організм з фенотипом\(X\) виживе до розмноження\(w(X)\), є, і ми будемо думати про це як про придатність нашого організму. Імовірність розподілу фенотипів у тих, хто виживає до розмноження, становить
\[\mathbb{P}(X | \textrm{survive}) = \frac{p(x) w(x)}{ \int_{-\infty}^{\infty} p(x) w(x) dx}.\]
де знаменник - це константа нормалізації, яка гарантує, що наш фенотипічний розподіл інтегрується в одне ціле. Знаменник також має тлумачення того, що це середня придатність населення, яку ми будемо називати\(\overline{w}\), тобто
\[\overline{w} = \int_{-\infty}^{\infty} p(x) w(x) dx. \label{eqn:pheno_mean_fitness}\]
Тому ми можемо написати середній фенотип у тих, хто виживає, щоб розмножуватися як
\[\mu_S = \frac{1}{\overline{w}}\int_{-\infty}^{\infty} x p(x) w(x) dx\]
Якщо мати на увазі центр розподілу фенотипів в нашій популяції, тобто встановити фенотип перед відбором на нуль, то
\[S=\mu_S= \frac{1}{\overline{w}}\int_{-\infty}^{\infty} x p(x) w(x) dx = \frac{1}{\overline{w}}\E \left (X w(X) \right)\]
де заключна частина випливає з того, що інтеграл приймає середнє\(X w(X)\) над населенням.
Оскільки наш фенотип є середнім центром (\(\mathbb{E}(X)=0\)), ми можемо бачити, що\(S\) має форму коваріації між нашим фенотипом\(X\) та нашою відносною придатністю\(\frac{w(X)}{\overline{w}}\).
\[S = \E \left (X \frac{w(X)}{\overline{w}} \right) =Cov \left(X, \frac{w(X)}{\overline{w}} \right) \label{S_covar}\]
Таким чином, наша зміна середнього фенотипу безпосередньо є мірою коваріації нашого фенотипу та нашої придатності. Переписуючи рівняння нашого селекціонера, використовуючи це спостереження, ми бачимо
\[R = \frac{V_A}{V_P} Cov \left(X, \frac{w(X)}{\overline{w}} \right)\]
ми бачимо, що реакція на відбір пов'язана з тим, що наша придатність (життєздатність) наших організмів/батьків збігається з нашим фенотипом, і що фенотип нашої дитини збігається з фенотипом нашого батька.
Фітнес-градієнти та лінійні регресії
Щоб зрозуміти це більш детально, давайте уявимо, що ми обчислюємо лінійну регресію середньоцентричного фенотипу\(i\) індивіда (\(X_i\)) на fitness (\(W_i\)), тобто
\[W_i \sim \beta X_i + \overline{w} \label{fitness_regression}\]
найкращий нахил цієї регресії (\(\beta\)), ми будемо називати його «градієнтом фітнесу», задається
\[\beta = Cov(X, \frac{w(X)}{\overline{w}} )/ V_P \label{beta_covar}\]
тобто фітнес-градієнт - це коваріація фенотип-фітнес, розділена на фенотипічну дисперсію. Використовуючи цей результат, ми можемо переписати рівняння селекціонера як
\[R= V_A \beta \label{eqn:R_beta}\]
тобто ми побачимо спрямовану відповідь на відбір, якщо існує лінійна залежність фенотипу від фітнесу, і якщо є адитивна генетична дисперсія для фенотипу. Як один із прикладів фітнес-градієнта, на малюнку\ ref {fig:red_deer_fitness_grad} життєвий репродуктивний успіх (LRS) чоловічого Red Deer побудовано проти ваги їх рогів. Червона лінія дає лінійну регресію фітнесу (LRS) на роги маси і нахил цієї лінії є фітнес-градієнт (\(\beta\)).

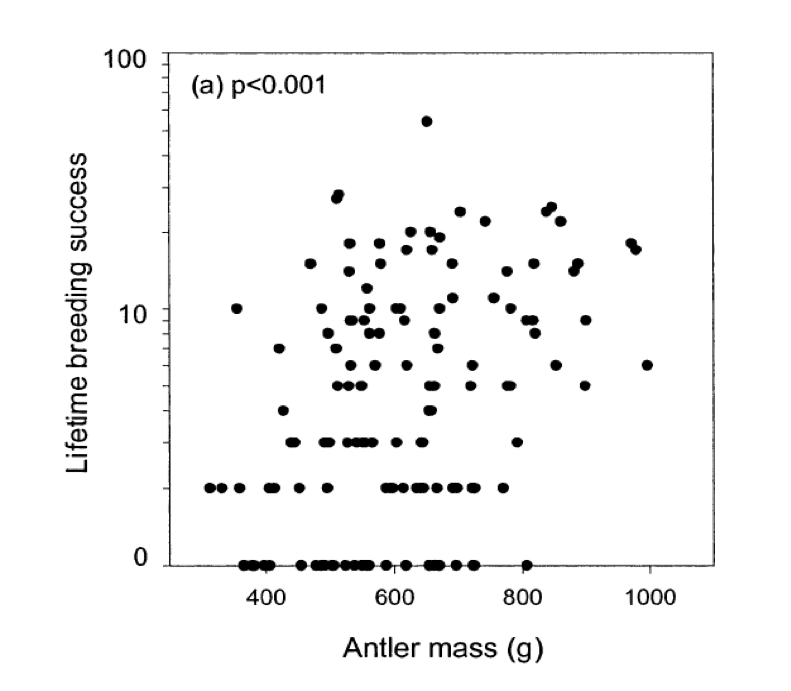
див. статтю для обговорення складнощів прирівнювання цього градієнта відбору до еволюційної відповіді. \ код коду {https://github.com/cooplab/popgen-no...n_grad_deer.R}.
Фундаментальна теорема Фішера про природний відбір
Нарешті, як розвивається середня придатність нашого населення? Якщо ми виберемо відносну придатність, щоб бути нашим фенотипом (\(X=\frac{w(X)}{\overline{w}}\)), то відповідь у фітнесі
\[\begin{aligned} R &= \frac{V_A}{V_P} Cov \left(\frac{w(X)}{\overline{w}} , \frac{w(X)}{\overline{w}} \right) = \frac{V_A}{V_P} V_P \nonumber\\ &=V_A\end{aligned}\]
тобто реакція на відбір дорівнює адитивної генетичної дисперсії для відносної придатності. Або як висловився Фішер
«Швидкість підвищення придатності будь-якого організму в будь-який час дорівнює його генетичній дисперсії в фітнесі в той час». - (pg 37)
Фішер назвав це «фундаментальною теоремою природного відбору». Нашим доказом тут є лише ескіз, і потрібні більш формальні підходи, щоб показати його в загальному вигляді. Там було багато скрегіт зубів над тим, наскільки широко цей результат тримає, і саме те, що Фішер мав на увазі.
Спрямований відбір на фітнес-ландшафти
Однією з поширених метафор, коли ми говоримо про еволюцію, є те, що населення досліджує адаптивний ландшафт з природним відбором, що штовхає населення до більш високих станів придатності, що відповідають пікам у цьому ландшафті (див. Рис.\ ref {Fig:Fitness_landscape_1d}). знайдено викликає формулювання Рівняння селекціонера, яке допомагає нашій інтуїції фенотипічних фітнес-ландшафтів. показало, що, якщо фенотип нормально розподілений, відповідь на виділення (\(R\)) може бути записана термінами градієнта (похідної\(\overline{w}\)) середньої придатності () населення як функція середнього фенотипу:
\[R = \frac{V_A}{\overline{w}} \frac{\partial \overline{w}}{\partial \bar{x}} \label{eqn:pheno_fitness_landscape} %V_A % \frac{\partial \log \left(\overline{w} \right)}{\partial \bar{z}}\]
Що це означає? Ну\(\frac{V_A}{\overline{w}}\) завжди позитивний, тому напрямок реагування нашої сукупності на відбір прогнозується за ознакою похідної (докладніше про похідні див. Розділ\ ref {sect:calculus}). Якщо збільшення середнього фенотипу популяції трохи збільшить середню придатність (\(\frac{\partial \overline{w}}{\partial \bar{x}} >0\)) наша популяція відповість на це покоління, еволюціонуючи до більш високих значень ознаки (\(R>0\)), ліва панель Figure\ ref {Fig:Fitness_landscape_1d_w_wBar}. І навпаки, якщо зменшення популяції середнього фенотипу трохи збільшить середню придатність (\(\frac{\partial \overline{w}}{\partial \bar{x}} <0\)), популяція буде розвиватися до нижчих значень фенотипу (середня панель Figure\ ref {fig:fitness_landscape_1d_w_wbar}). Таким чином, якщо тиск на відбір залишається постійним, ми можемо думати про населення як еволюціонує на адаптивному ландшафті, де висота надається населенням середньої придатності. Природний відбір діє на основі індивідуальної фізичної підготовки, але в результаті цього наше населення збільшується в своїй середній придатності, тобто наше населення стає краще пристосованим. Нижче ми обговоримо застереження цієї інтерпретації сходження на гору.
Що відбувається, коли він досягає вершини піку? Добре на вершині піку\(\frac{\partial \overline{w}}{\partial \bar{x}}=0\), так як це локальний максимум, і так\(R=0\). Припускаючи, що зв'язок між фітнесом і фенотипом залишається постійним, наше населення залишиться на вершині фітнес-піку. Такий погляд на природний відбір не означає, що населення розвивається до найкращого стану. Наше населення просто йде вгору по пагорбу середнього фітнесу (кінцева панель Малюнок\ ref {Fig:Fitness_landscape_1D_W_WBar}). Однак цей пік не обов'язково найвищий фітнес-пік, а просто незалежно від того, який пік був найближчим. Таким чином, наше населення може потрапити в пастку на локальний, але не глобальний пік фітнесу (див., наприклад, рис.\ ref {Fig:Fitness_landscape_1d}).
Один драматичний приклад, що документує адаптивну еволюцію до нового оптимального фітнесу, пропонується чудовою часовою серією еволюції з викопного озера в Неваді. У цьому озері шари осаду закладені щороку, що дозволяє дуже детальний часовий ряд з більш ніж п'ятьма тисячами скам'янілостей виміряних. Часовий ряд документує еволюцію до нового набору оптимальних фенотипів у п'ятнадцять тисяч років після початкового вторгнення в озеро сильно броньованих видів колючок. На малюнку\ ref {Fig:Stickleback_fossil_traj} популяція середня кількість торкаються птеригіофорів, кісток, що підтримують спинні шипи, через викопний запис (рис.\ ref {Fig:Stickleback_fossil}). Зверніть увагу, як швидко вид розвивається до своєї нової цінності, імовірно, оптимальної придатності в їх новому середовищі, і довгий наступний проміжок часу, протягом якого популяція означає фенотип коливається щодо свого нового значення.
підігнана модель населення, що адаптується до фітнес-ландшафту, з єдиним піком, до цих даних часових рядів. Їх пристосована поверхня показана на нижній панелі малюнка\ ref {fig:stickleback_fossil_traj}. Стрілки показують ходи, які населення означає фенотип робить на цій передбачуваній поверхні придатності. Населення спочатку робить великі кроки вгору до піку цієї поверхні і згодом коливається навколо піку. За інтерпретацією, що існує єдиний стаціонарний пік, ці коливання представляють генетичний дрейф, випадковим чином збиваючи популяцію з її оптимального, з відбором, що діє для відновлення популяції до цього локального оптимального.
Проблеми з інтерпретацією фітнес-пейзажів.
На практиці фітнес-пейзажі можуть бути не постійними. Навколишнє середовище може постійно змінюватися, тому наше населення постійно змушене змінюватися, щоб не відставати від піку фітнесу. Дійсно, наше середовище може змінитися так швидко, що наше населення не може йти в ногу з піком. Наше населення все ще намагається збільшити свою середню придатність, «адаптуватися», але сам ландшафт розвивається. У разі дуже швидких змін навколишнього середовища наше населення може скочуватися все далі і далі від піку, і, як наслідок, його середня придатність зменшується, що може призвести населення до вимирання, якщо наше населення опуститься нижче досить\(\overline{w}<1\) довго. Умови вимирання є активним напрямком досліджень у галузі «Еволюційного порятунку». Більш загально, для того, щоб наш результат фітнес-ландшафту (eqn\ ref {eqn:pheno_fitness_landscape}) тримався, і для того, щоб ми могли говорити про наше населення, яке намагається еволюціонувати до вищих середніх станів фітнесу, нам потрібна придатність наших фенотипів, щоб бути незалежними від частоти інших фенотипів у популяції. (Ця незалежність дозволяє припустити, що придатність індивідів не є функцією середнього фенотипу, як це потрібно в eqn\ ref {eqn:proof_landscape}). Припущення про частотну незалежність може не триматися, коли існує конкуренція між окремими особами, наприклад, за ресурси або товариші, оскільки тоді придатність індивіда залежить від стратегій, які проводять інші особи в популяціях.
Стабілізуючий та руйнівний відбір
До цих пір ми щойно розглядали спрямований відбір, де відбір діє, щоб змінити середній фенотип. Однак ми також можемо використовувати кількісні генетичні моделі для опису інших способів відбору, виходячи з впливу на популяцію означає, що наступним природним кроком є думка про відбір, який діє на дисперсію популяції. Вибір може діяти сильніше проти осіб у хвостах розподілу, причому ті, хто ближче до середнього фенотипу, мають вищу придатність, що знижує дисперсію. Вибір може також зневажати особам, близьким до середнього населення, з особами з екстремальними фенотипами, що мають вищу придатність, що діє для збільшення дисперсії населення.
Спрямований відбір відбувається через коваріацію між нашим фенотипом та фітнесом, eqn\ ref {S_CoVar}. Подібно до того, як вираження спрямованого відбору як коваріації дозволило нам охарактеризувати спрямований відбір як лінійну залежність між фітнесом та фенотипом\(\beta\), ми можемо узагальнити вибір зменшення дисперсії, включивши квадратичний термін у регресію придатності на фенотип
\[w_i \sim \beta x_i + \frac{1}{2} \gamma x_i^2 + \overline{w} \label{fitness_regression_stab}\]
Це\(\gamma\), коефіцієнт квадратичного члена в нашій моделі, є квадратичним градієнтом відбору: коваріацією придатності та квадратичним відхиленням від фенотипічного середнього (\(\mu_{BS}\)), тобто
\[\gamma = \frac{Cov\left(w(X), (X-\mu_{BS})^2 \right)}{V^2}\]
Наша\(\gamma\) описує кривизну поверхні фітнесу навколо середнього. Значення\(\gamma<0\) узгоджуються зі стабілізуючим відбором, зменшуючи дисперсію. У той час як значення\(\gamma>0\) узгоджуються з руйнівним відбором, збільшуючи дисперсію.
При стабілізуючому відборі особи з екстремальними фенотипами в будь-якому хвості мають меншу придатність, результатом якої є зменшення фенотипової дисперсії в межах покоління. Класичним випадком стабілізації виділення є вага при народженні у людини. Мері Карн зібрала дані про майже чотирнадцять тисяч вагітностей з 1935-46 років щодо ваги при народженні та смертності. Ці дані передаються на рисунку\ ref {рис:birth_weight}. Дисперсія всіх пологів становить\(1.575\) lb\(^2\), тоді як у живих пологів це було зменшено до\(1.26\) lb\(^2\), зменшення дисперсії на 20% через стабілізуючий відбір. Варто зазначити, що цей тиск на відбір значно зменшився протягом десятиліть у суспільствах, які мають доступ до хорошої пренатальної допомоги.
У Центральній Африці чорнобрюхие насіннєві крекери (Pyrenestes ostrinus) демонструють руйнівний відбір на чудовому поліморфізмі розміром з дзьоб (рис.\ ref {black_bellied_seedcrackers_beaks}). Дрібнодзьобові особини харчуються м'яким насінням з одного виду болотної осоки, тоді як великодзьобові особини харчуються твердими насінням з іншої осоки, що вимагає в десять разів більшої сили для розтріскування. зафіксували долі сотень молодих, і виявили, що особини з проміжними розмірами дзьоба виживали набагато нижчі показники (рис.\ ref {black_bellied_seedCrackers_beaks}), оскільки вони не були добре адаптовані до будь-якого ресурсу насіння. Довжина розриву підлягає руйнівному відбору, як це також можна побачити за значним негативним квадратичним терміном у регресії ймовірності виживання на довжині розриву. Дисперсія довжини нижньої щелепи в загальній вибірці особин становила\(0.5\) мм,\(^2\) у тих, хто вижив, ця дисперсія збільшувалася\(2.5\) в рази до\(1.3\) мм\(^2\).
Щоб проілюструвати, як спрямований відбір та квадратичні терміни відіграють під час адаптації, розглянемо жовчну муху золотарника (Eurosta solidaginis), він же золотарник м'яча. Див. Рисунок\ ref {gall_size_stab}. Як випливає з чудової назви, ця комаха відкладає свої яйця в рослини золотарника, а личинки виділяють хімічні речовини, змушуючи рослину утворювати жовч, який утворює будинок для личинок у міру їх розвитку. Хоча це здається досить солодкою угодою для личинок, це не без його небезпеки.
Коли маленькі, кулькові галли стають жертвою паразитизму від паразитоїдних ос. Коли всі кульові галли невеликі, у популяції відбір призводить сильний позитивний спрямований відбір на розмір жовчного, з невеликим стабілізуючим відбором. Зверніть увагу на ліву панель Figure\ ref {gall_size_stab} хорошу згоду між лінійним градієнтом вибору та підгонкою, включаючи лінійний та квадратичний термін. Однак більші кульки потрапляють під палець хижацтва від пухових дятлів і чорношапкових птахів, які шукають смачних личинок. Таким чином, проміжні розміри галли віддають перевагу, пік фітнесу, який населення швидко досягає. Одного разу на цьому піку, як показано на правій панелі Figure\ ref {gall_size_stab}, немає вибору спрямованості, тобто немає лінійного нахилу, але є сильне стабілізуюче виділення, тобто квадратичний термін. Таким чином, населення буде підтримуватися на цьому піку фітнесу нескінченно довго, якщо навколишнє середовище залишиться незмінним.
Фенотипічний природний відбір вимагає варіації фенотипів впливає на фітнес невипадковим чином. Для еволюції шляхом природного відбору нам потрібно, щоб ця фенотипічна варіація була спадковою.
Проста модель реакції на спрямований відбір на фенотип наведено рівнянням селекціонера. Очікувана реакція на вибір між суміжними поколіннями (\(R\)) пропорційна реакції на виділення в межах покоління, диференціала вибору (\(S\)), де константа пропорційності є вузькочущою спадковістю (\(h^2\)) риса. Таким чином, ми очікуємо побачити і дійсно бачимо сильну реакцію на відбір, коли селекція спричиняє великі зміни в поколінні спадкових фенотипів.
Якщо тиск відбору та спадковість залишаються постійними, ми очікуємо лінійної реакції на вибір у багатьох поколіннях. Ми часто можемо бачити це в селекційних експериментах, але в дикій природі тиск відбору часто коливається від покоління до покоління. Великі зміни фенотипу, які ми бачимо в записі викопних копалин, легко пояснюються сильними сторонами відбору, які ми бачимо, діють протягом коротких часових шкал.
Є ще два поширені способи написання рівняння селекціонера. Перший використовує градієнт виділення (\(\beta\)), регресія фітнесу за фенотипом. По-друге, інтерпретація фітнес-ландшафту спирається на написання цього градієнта вибору як похідного від середньої придатності за фенотипом. Форма фітнес-ландшафту до рівняння заводчика допомагає нам зрозуміти, як і коли ми можемо очікувати, що відбір діятиме, щоб збільшити середню придатність населення.
Ми можемо зрозуміти інші форми відбору на нашому фенотипі, які, як очікується, будуть діяти на фенотипічну дисперсію, а не на середнє, наприклад, руйнівний та стабілізуючий відбір, шляхом розширення рівняння селекціонера, щоб включити терміни для квадратичних градієнтів відбору.
Ви вивчаєте швидку еволюцію розміру легких органів у світлячків (Photinus pyralis) у відповідь на світлове забруднення на прерії в Огайо. У січні 1985 року через прерії було побудовано шосе з яскравими вуличними ліхтарями. Оскільки світлячки використовують світлові сигнали для знаходження товаришів, особин з меншими, а отже, і менш видимими, легкі органи були менш успішними при спаровуванні в цих нових умовах освітлення. Ви знаєте, що легкий орган був, в середньому, завдовжки 4 мм до будівництва шосе. У 2005 році середній розмір легких органів у цієї популяції до спарювання становив 6 мм. Якщо цей світлячок має 1 покоління на рік, а успадкованість вузького смислу становить 0,1, якою була середня довжина світлового органу успішно розмножуються особин у 1985 році (перший рік селекції)?
Ви заводчик кроликів, і ви вирішуєте, що хочете кроликів з довгим хутром. Фенотипічна дисперсія є\(4\textrm{cm}^2\). Коваріація довжини хутра між повними сибами є\(1\textrm{cm}^2\). Середня довжина хутра у початкової популяції становить\(3\textrm{cm}\). Ви вирішите розводити верхівку\(\frac{1}{3}\) популяції найдовшим хутром, а їх середня довжина хутра -\(5\) см.
Припускаючи, що коваріація між сибами обумовлена лише адитивною генетичною дисперсією, скільки поколінь цього режиму відбору знадобиться, щоб довжина хутра була\(10cm\) в популяції?