2.3: Структура та функція - Білки I
- Page ID
- 2491

Джерело: BiochemFFA_2_2.pdf. Весь підручник доступний безкоштовно від авторів за адресою http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
Білки - це робочі конячки клітини. Практично все, що відбувається всередині клітин, відбувається в результаті дії білків. Крім усього іншого, білкові ферменти каталізують переважну більшість клітинних реакцій, опосередковують сигналізацію, надають структуру як клітинам, так і багатоклітинним організмам, здійснюють контроль над експресією генів. Життя, як ми її знаємо, не існувало б, якби не було білків. Універсальність білків виникає через їх різноманітної структури.
Білки виробляються шляхом зв'язування між собою амінокислот, причому кожен білок має характерну і унікальну амінокислотну послідовність. Щоб отримати відчуття різноманітності білків, які можуть бути зроблені з використанням 20 різних амінокислот, врахуйте, що кількість різних комбінацій, можливих з 20 амінокислотами, становить 20 n, де n=кількість амінокислот в ланцюжку. Стає очевидним, що навіть дипептид, виготовлений лише з двох амінокислот, з'єднаних разом, дає нам 20 2 = 400 різних комбінацій. Якщо ми зробимо розрахунок для короткого пептиду з 10 амінокислот, ми отримаємо величезні 10 240 000 000 комбінацій. Більшість білків набагато більше, ніж це, що робить можливу кількість білків з унікальними амінокислотними послідовностями неймовірно величезним.
Рівні структури
Значення унікальної послідовності, або порядку, амінокислот, відомих як первинна структура білка, полягає в тому, що він диктує 3-D конформацію складеного білка буде мати. Ця конформація, в свою чергу, визначить функцію білка. Розглянемо білкову структуру на чотирьох різних рівнях (рис. 2.17) - 1) як послідовність амінокислот в білку (первинна структура) дає ідентичність і характеристики білка (рис. 2.18); 2) як локальні взаємодії між однією частиною поліпептидного хребта та іншою впливають на форму білка ( вторинна структура); 3) як поліпептидний ланцюг білка може складатися, щоб дозволити амінокислотам взаємодіяти один з одним, які не близькі за первинною структурою (третинної структури); і 4) як різні поліпептидні ланцюги взаємодіють один з одним в межах багатосубодиничного білка (четвертинна структура).
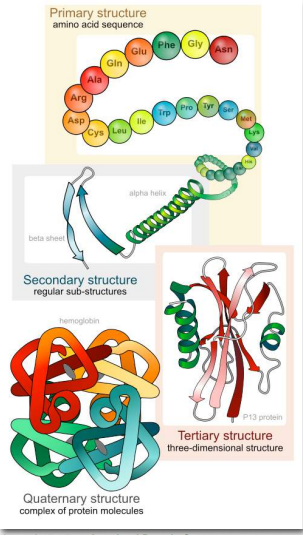
На цьому етапі ми повинні надати пару визначень. Ми використовуємо термін поліпептид для позначення одного полімеру амінокислот. Вона може, а може і не склалася в остаточну, функціональну форму. Термін білок іноді використовується як взаємозамінний з поліпептидом, як у «синтезі білка». Однак, як правило, використовується для позначення складеної функціональної молекули, яка може мати одну або кілька субодиниць (що складаються з окремих поліпептидів). Таким чином, коли ми використовуємо термін білок, ми зазвичай маємо на увазі функціональний, складчастий поліпептид або пептиди. Структура має важливе значення для функції. Якщо ви змінюєте структуру, ви змінюєте функцію - зазвичай, але не завжди, це означає, що ви втратите всю функцію. Для багатьох білків змінити структуру нескладно.
Білки гнучкі, не жорстко закріплені за структурою. Як ми побачимо, саме гнучкість білків дозволяє їм бути дивовижними каталізаторами і дозволяє їм адаптуватися, реагувати і передавати сигнали при зв'язуванні інших молекул або білків. Однак білки не є нескінченно гнучкими. Існують обмеження на конформації, які можуть прийняти білки, і ці обмеження регулюють конформації, які відображають білки.
Тонкі зміни
Навіть дуже крихітні, тонкі зміни в структурі білка можуть спричинити великі зміни в поведінці білків. Наприклад, гемоглобін зазнає неймовірно невеликих структурних змін при зв'язуванні однієї молекули кисню, і ця проста зміна призводить до того, що решта білка отримує значно більшу спорідненість до кисню, якого білок не мав до структурних змін.
Послідовність, структура і функції
Як обговорювалося раніше, кількість різних амінокислотних послідовностей можливих, навіть для коротких пептидів, дуже велика. Немає двох білків з різними амінокислотними послідовностями (первинна структура) не мають однакової загальної структури. Унікальна амінокислотна послідовність білка відбивається в його унікальній складчастої структурі. Ця структура, в свою чергу, визначає функцію білка. Ось чому мутації, які змінюють послідовність амінокислот, можуть впливати на функцію білка.
Синтез білка
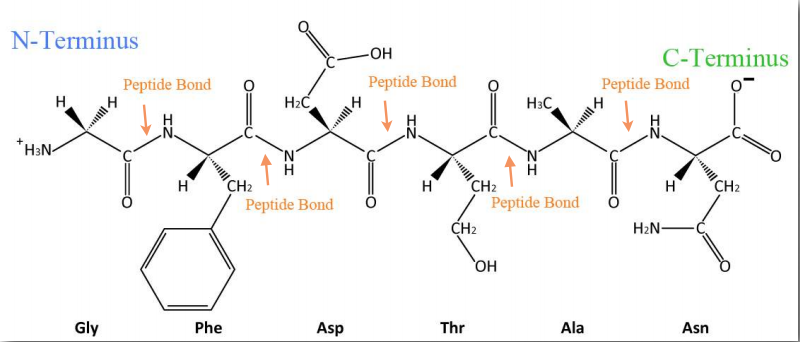
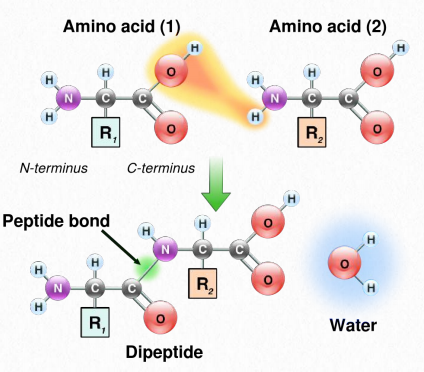
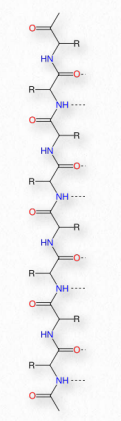
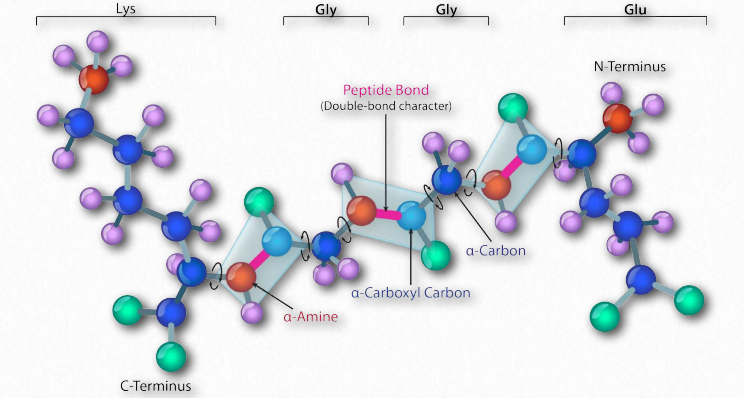
Синтез білків відбувається в рибосомах і протікає шляхом приєднання карбоксильного кінця першої амінокислоти до амінокінця наступної (рис. 2.19). Кінець білка, який має вільну α-аміногрупу, називається амінокінцевим або N-кінцевим. Інший кінець називається карбоксильним кінцем або C-кінцевим, так як він містить єдину вільну α-карбоксильну групу. Всі інші α-аміногрупи та α-карбоксильні групи пов'язані в утворенні пептиду Рис. 2.19 Зв'язування амінокислот через зв'язки формування пептидних зв'язків, які з'єднують сусідні амінокислоти разом. Протеїни синтезуються, починаючи з амінокінцевого і закінчуючи на карбоксильному кінці.
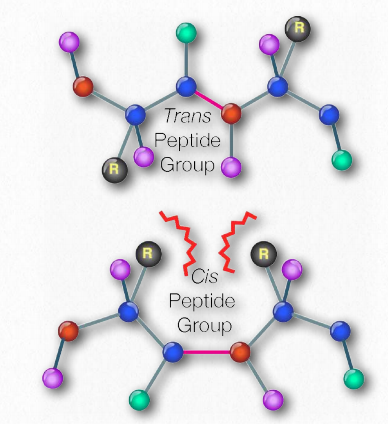
Схематично на малюнку 2.18 ми можемо побачити, як послідовні R-групи білка розташовані в змінній орієнтації по обидва боки поліпептидного ланцюга. Організація R-груп таким чином не випадкова. Стерична перешкода може виникати при орієнтації послідовних R-груп на одній стороні пептидного хребта (рис. 2.20)
Первинна структура
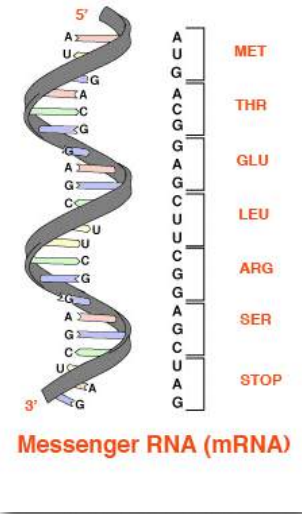
Первинна структура є кінцевою детермінантою загальної конформації білка. Первинна структура будь-якого білка прибула в свій поточний стан в результаті мутації і відбору протягом еволюційного часу. Первинна структура білків закріплена послідовністю кодування ДНК для нього в геномі. Регіони ДНК, що визначають білки, відомі як кодуючі області (або гени).
Базові послідовності цих областей безпосередньо визначають послідовність амінокислот в білках, з відповідністю один до одного між кодонами (групами з трьох послідовних основ) в ДНК і амінокислотами в закодованому білку. Послідовність кодонів в ДНК, скопійована в месенджерну РНК, визначає послідовність амінокислот в білку. (Малюнок 2.21).
Порядок, в якому амінокислоти з'єднуються разом в синтезі білка починає визначати набір взаємодій між амінокислотами, навіть коли відбувається синтез. Тобто поліпептид може складатися навіть у міру його виготовлення. Порядок структур R-групи та результуючі взаємодії дуже важливі, оскільки ранні взаємодії впливають на пізніші взаємодії. Це пояснюється тим, що взаємодії починають встановлювати структури - вторинні та третинні. Якщо, наприклад, починає формуватися гвинтова структура (вторинна структура), то можливості взаємодії тієї чи іншої амінокислоти Rгрупи можуть бути іншими, ніж якби спіраль не утворилася (рис. 2.22). Взаємодія R-групи також може спричинити вигини поліпептидної послідовності (третинної структури), і ці вигини можуть створювати (в деяких випадках) можливості для взаємодій, які були б неможливі без вигину або запобігти (в інших випадках) подібні можливості взаємодії.
вторинна структура
У міру прогресування синтезу білка починають відбуватися взаємодії між амінокислотами, близькими один до одного, що породжує місцеві закономірності, звані вторинною структурою. До таких вторинних структур відносяться добре відомі α- спіраль і β-нитки. Обидва були передбачені Лінусом Полінгом, Робертом Корі і Германом Бренсоном в 1951 році. Кожна споруда має унікальні особливості.
α-спіраль
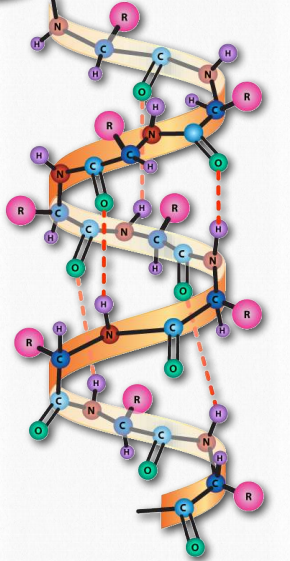
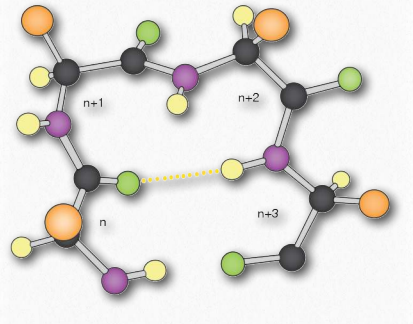
α-спіраль має згорнуту структуру, з 3,6 амінокислотами на оборот спіралі (5 спіральних витків = 18 амінокислот). Спіралі переважно правші - лише в рідкісних випадках, наприклад, в послідовності з багатьма гліцинами можуть утворюватися ліві α- спіралі. У α-спіралі водневі зв'язки утворюються між групами C = O та N-H групами в поліпептидному хребті, які є чотирма віддаленими амінокислотами. Ці водневі зв'язки є первинними силами, що стабілізують α-спіраль.
Ми використовуємо терміни підйом, повторення та крок для опису параметрів будь-якої спіралі. Повторення - це кількість залишків у спіралі, перш ніж вона почне повторюватися. Для α-спіралі повторення становить 3,6 амінокислоти на оборот спіралі. Підйом - це відстань, яку спираль піднімає з додаванням кожного залишку. Для α-спіралі це 0,15 нм на амінокислоту. Крок - це відстань між повними витками спіралі. Для α-спіралі це 0,54 нм. Стійкість α-спіралі посилюється наявністю амінокислоти аспартату.

β нитка/лист
Спіраль - це, звичайно ж, тривимірний об'єкт. Сплющена форма спіралі в двох вимірах є загальним описом для β- пасма. Замість котушок, β-нитки мають вигини, і їх іноді називають складками, як складки в завісі. β-нитки можуть бути організовані для формування ретельно організованих структур, таких як листи, бочки та інші механізми.
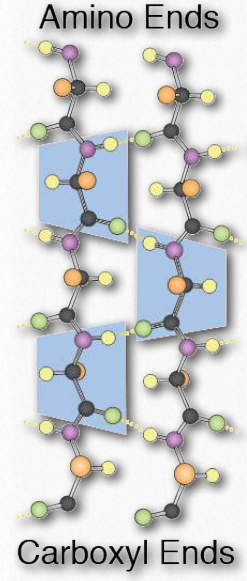
Структури β-нитки вищого порядку іноді називають надвторинними структурами), оскільки вони передбачають взаємодію між амінокислотами, не близькими в первинній послідовності. Ці структури теж стабілізуються водневими зв'язками між атомами карбонільного кисню і воднями амінних груп в поліпептидному хребті (рис. 2.28). У структурі вищого порядку пасма можуть розташовуватися паралельно (амінокарбоксильні орієнтації однакові) або антипаралельні (амінокарбоксильні орієнтації навпроти один одного (на малюнку 2.27 напрямок пасма показано стрілкою на діаграмах стрічки).
Повороти
Повороти (іноді їх називають зворотними поворотами) - це тип вторинної структури, яка, як випливає з назви, викликає поворот в структурі поліпептидного ланцюга. Повороти породжують третинну структуру в кінцевому підсумку, викликаючи перебої у вторинних структурах (α- спіралі та β-нитки) і часто служать сполучними областями між двома областями вторинної структури в білку. Пролін і гліцин відіграють загальні ролі по черзі, забезпечуючи меншу гнучкість (починаючи поворот) і більшу гнучкість (полегшуючи поворот) відповідно.
Існує щонайменше п'ять типів поворотів, з численними варіаціями кожного з яких виникає безліч різних поворотів. П'ять типів поворотів:
• Δ-витки - кінцеві амінокислоти розділені однією пептидною зв'язком
• γ-витки - поділ двома пептидними зв'язками
•β-витки - поділ трьома пептидними зв'язками
•α-витки - поділ чотирма пептидними зв'язками
•π-витки - поділ на п'ять зв'язків
З них β-витки є найбільш поширеною формою, а Δ-витки - теоретичними, але малоймовірними, через стеричні обмеження. На малюнку 2.29 зображено β- поворот.
3 10 спіралей
Крім α-спіралі, β-ниток і різних поворотів, в білках спостерігаються інші регулярні, повторювані структури, але зустрічаються набагато рідше. Спіраль 3 10 є четвертою найбільш поширеною вторинною структурою в білках, що становить близько 10-15% всіх спіралей. Спіраль отримала свою назву від того, що вона містить 10 амінокислот в 3 обороти. Це правша. Водневі зв'язки утворюються між амінокислотами, які є трьома залишками один від одного Найчастіше спіраль 3 10 з'являється на амінному або карбоксильному кінці α-спіралі. Як і α-спіраль, спіраль 3 10 стабілізується наявністю аспартату в її послідовності.
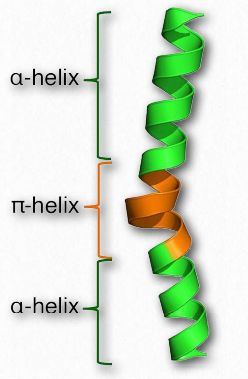
π-спіралі
π-спіраль може розглядатися як особливий тип α- спіралі. Деякі джерела описують його як α-спіраль з зайвою амінокислотою, застрягла в середині неї (рис. 2.32). π-спіралі не зовсім рідкісні, зустрічаються хоча б раз в цілих 15% всіх білків. Як і α- спіраль, π-спіраль правостороння, але там, де α-спіраль має 18 амінокислот в 5 оборотах, π-спіраль має 22 амінокислоти в 5 оборотах. π-спіралі зазвичай не розтягуються на дуже великі відстані. Більшість з них тільки близько 7 амінокислот довго і послідовність майже завжди відбувається в середині α-спіральної області.
Рамачандран ділянки
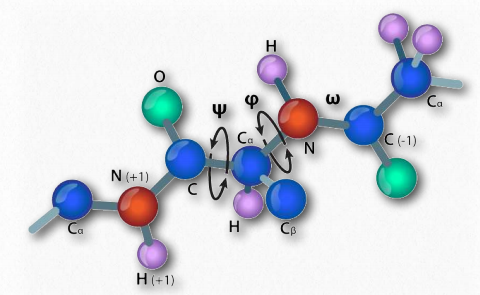
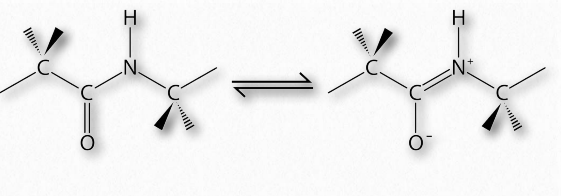
У 1963 році Г.Н. Рамачандран, Рамакрішнан і В.Сасісехаран описали новий спосіб опису структури білка. Якщо розглядати кістяк поліпептидного ланцюга, то він складається з повторюваного набору з трьох зв'язків. Послідовно (в амінокарбоксильному напрямку) вони являють собою 1) обертається зв'язок (ψ) між α-вуглецем і α-карбоксилом, що передує пептидному зв'язку (див. ТУТ), 2) не обертається пептидний зв'язок (ω) між α-карбоксильною і α-амінними групами) і 3) обертається зв'язок (φ) між α-аміном і α-вуглецем після пептидний зв'язок (див. ТУТ). Зверніть увагу на малюнках 2.33 та 2.34, що напрямок амінокарбоксилу знаходиться справа наліво.
Наявність карбонільного кисню на α-карбоксильній групі дозволяє пептидному зв'язку існувати як резонансна структура, що означає, що він поводиться деякий час як подвійний зв'язок. Подвійні облігації, звичайно, не можуть обертатися, але зв'язки по обидва боки від нього мають деяку свободу обертання. Кути φ і ψ обмежені певними значеннями, оскільки деякі кути призведуть до стеричної перешкоди. Крім того, кожен тип вторинної конструкції має характерний діапазон значень для φ і ψ.
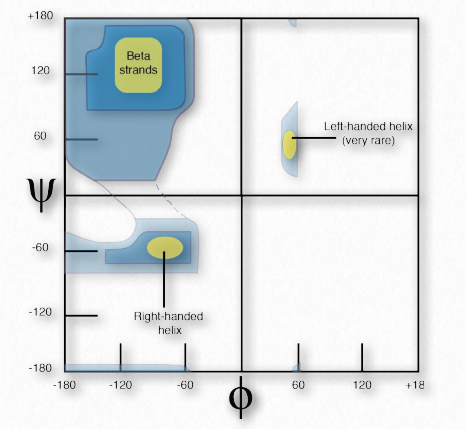
Рамачандран та його колеги провели теоретичні розрахунки енергетичної стійкості всіх можливих кутів від 0° до 360° для кожного з φ і ψ кутів і побудували результати на графіку Рамачандрана (також званий ділянкою φ-ψ), окресливши області кутів, які теоретично були найбільш стійкими (рис. 2.35).
Виявлено три первинні області стійкості, що відповідають φ-ψ кутам β-ниток (вгорі ліворуч), праворучним α- спіралям (знизу ліворуч) та лівим α-спіралям (вгорі праворуч). Графіки прогнозованої стійкості надзвичайно точні у порівнянні з φ-ψ кутами фактичних білків.
Передбачення вторинної структури
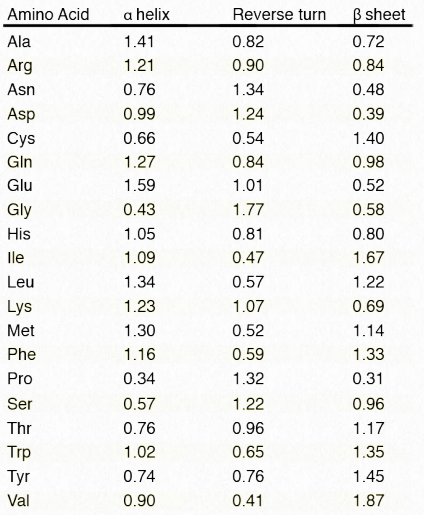
Таблиця 2.3 - Відносні тенденції кожної амінокислоти перебувати у вторинній структурі. Більш високі значення вказують на більшу тенденцію Зображення Пенелопи Ірвінг
Порівнюючи первинну структуру (послідовності амінокислот) з відомими 3D-білковими структурами, можна підрахувати кожен раз, коли амінокислота виявляється в α-спіралі, β-пасмі/аркуші або повороті. Комп'ютерний аналіз тисяч цих послідовностей дозволяє призначити ймовірність появи будь-якої заданої амінокислоти в кожній з цих структур. Використовуючи ці тенденції, можна з точністю до 80% прогнозувати області вторинної структури в білку на основі виключно амінокислотної послідовності.
Це видно в таблиці 2.3. Виникнення в первинній послідовності трьох послідовних амінокислот з відносними тенденціями вище одиниці є показником того, що ця область поліпептиду знаходиться у відповідній вторинній структурі. Онлайн-ресурс для прогнозування вторинних структур під назвою PSIPRED доступний ТУТ.
гідрофобність
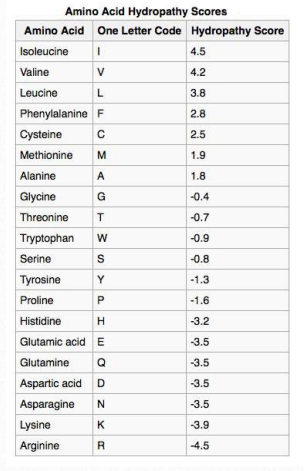
Таблиця 2.4 - Оцінка гідропатії
Хімія амінокислот Rgroups впливає на структури, в яких вони найчастіше зустрічаються. Підмножини їх хімічних властивостей можуть дати підказки до структури, а іноді і клітинного розташування. Яскравим прикладом є гідрофобність (тенденції уникнення води) деяких Rгруп. З огляду на водне середовище клітини, такі R-групи навряд чи виявляться на зовнішній поверхні згорнутого білка.
Однак це правило не діє для областей білка, які можуть бути вбудовані в ліпідні бішари клітинних/органельних мембран. Це пояснюється тим, що область таких білків, що утворюють трансмембранні домени, поховані в гідрофобному середовищі посередині ліпідного бішару.
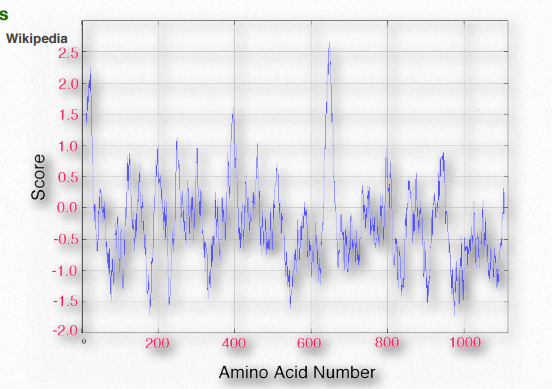
Не дивно, що сканування первинних послідовностей для конкретно розмірних/віддалених ділянок гідрофобних амінокислот може допомогти ідентифікувати білки, знайдені в мембранах. У таблиці 2.4 наведені значення гідрофобності для R-груп амінокислот. У цьому наборі шкала проходить від позитивних значень (гідрофобних) до від'ємних (гідрофільних). Графік гідропатії KytedooLittle для протеїну мембрани протоонкогену RET показаний на малюнку 2.36. Дві області білка дуже гідрофобні, як видно з піків поблизу амінокислот 5-10 і 630-640. Такі регіони можуть бути обґрунтовано очікувані, що вони будуть розташовуватися або всередині складеного білка, або бути частиною трансмембранних доменів.
Випадкові котушки
Деякі ділянки білка не припускають регулярної, помітної структури і іноді кажуть, що вони не мають вторинної структури, хоча вони можуть мати водневі зв'язки. Такі сегменти описуються як перебувають у випадкових котушках і можуть мати плинність до своєї структури, що призводить до того, що вони мають кілька стабільних форм. Випадкові котушки ідентифікуються за допомогою спектроскопічних методів, таких як круговий дихроїзм Вікіпедії та ядерний магнітний резонанс (ЯМР), в якому спостерігаються відмінні сигнали. Див. Також метаморфічні білки (ТУТ) та іскромно невпорядковані білки (ТУТ).

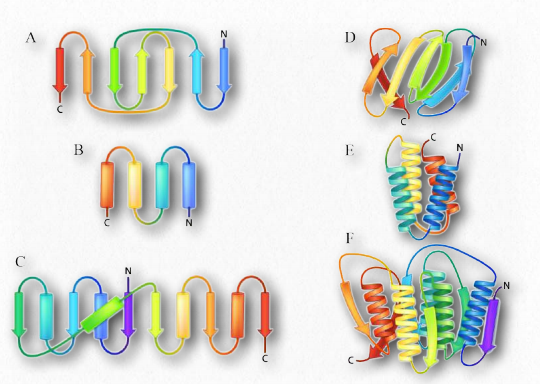
Надвторинна структура
Інший елемент білкової структури важче класифікувати, оскільки він включає в себе елементи вторинної та третинної структури. Названі надвторинною структурою (або структурними мотивами), ці структури містять кілька сусідніх компонентів вторинної структури, розташованих певним чином і які з'являються в декількох білках. Оскільки існує безліч способів виготовлення другорядних конструкцій з різних первинних структур, тому теж можуть виникати схожі мотиви з різних первинних послідовностей. Приклад структурного мотиву показаний на малюнку 2.37.
третинна структура
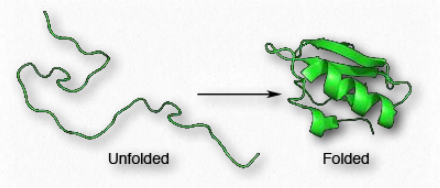
Білки відрізняються один від одного послідовністю містять їх амінокислот. Послідовність амінокислот білка визначає форму білка, оскільки хімічні властивості кожної амінокислоти - це сили, що породжують міжмолекулярні взаємодії, щоб почати створювати вторинні структури, такі як α-спіралі і β-нитки. Послідовність також визначає витки і випадкові котушки, які відіграють важливу роль в процесі згортання білка.
Оскільки форма має важливе значення для білкової функції, послідовність амінокислот породжує всі властивості білка. У міру протікання синтезу білка окремі компоненти вторинної структури починають взаємодіяти один з одним, породжуючи складки, які зближують між собою амінокислоти, які не знаходяться поруч один з одним в первинній структурі (рис. 2.38). На третинному рівні структури роль у згортанні відіграють взаємодії між R-групами амінокислот в білку, а також між поліпептидним хребтом і бічними групами амінокислот.
кулясті білки
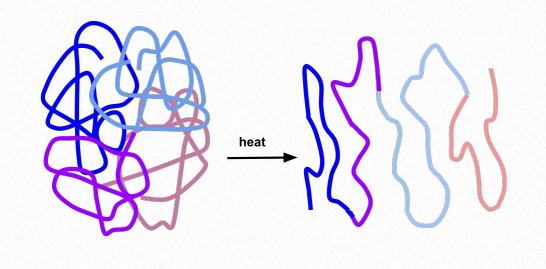
Складання породжує чіткі 3-D форми в білках, які не волокнисті. Ці білки називаються кулястими. Кульовий білок стабілізується тими ж силами, які керують його утворенням. До них відносяться іонні взаємодії, водневий зв'язок, гідрофобні сили, іонні зв'язки, дисульфідні зв'язки і металеві зв'язки. Такі процедури, як тепло, зміни рН, миючі засоби, сечовина та меркаптоетанол, долають стабілізуючі сили і змушують білок розгортатися, втрачаючи свою структуру і (зазвичай) свою функцію (рис. 2.39). Здатність тепла і миючих засобів денатурувати білки - ось чому ми готуємо їжу і миємо руки перед їжею - такі процедури денатурують білки в мікроорганізмах на наших руках. Організми, які живуть у середовищах з високою температурою (понад 50° C), мають білки зі зміною стабілізуючих сил - додаткові водневі зв'язки, додаткові сольові мости (іонні взаємодії), і компактність може відігравати роль у запобіганні розгортання цих білків.
Білкові стабілізуючі сили
Перш ніж розглядати процес згортання, розглянемо деякі сили, які допомагають стабілізувати білки.
водневі зв'язки
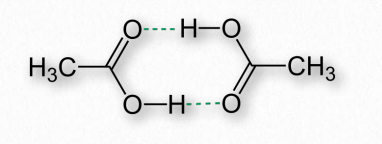
Водневі зв'язки виникають в результаті частково заряджених воднів, виявлених в ковалентних зв'язках. Це відбувається, коли атом, з яким пов'язаний водень, має більшу електронегативність, ніж сам водень, в результаті чого водень має частковий позитивний заряд, оскільки він не здатний утримувати електрони близько до себе (рис. 2.40).
Частково заряджений таким чином водень притягується до атомів, таких як кисень та азот, які мають часткові негативні заряди, завдяки більшій електронегативності і, таким чином, утримуючи електрони ближче до себе. Частково позитивно заряджені водні називають донорами, тоді як частково негативні атоми, до яких вони притягуються, називаються акцепторами. (Див. Малюнок 1.30).
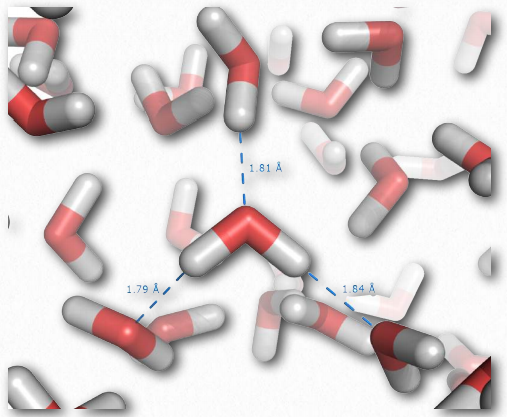
Окремі водневі зв'язки набагато слабкіше ковалентного зв'язку, але в сукупності вони можуть надавати сильні сили. Розглянемо рідку воду, яка містить величезну кількість водневих зв'язків (рис. 2.41). Ці сили допомагають воді залишатися рідкою при кімнатній температурі. Інші молекули, які не мають водневих зв'язків рівної або більшої молекулярної маси, ніж вода, такі як метан або вуглекислий газ, є газами з однаковою температурою. Таким чином, міжмолекулярні взаємодії між молекулами води допомагають «утримувати» воду разом і залишатися рідиною. Примітно, що лише підвищуючи температуру води до кипіння, сили водневого зв'язку подолані, дозволяючи воді стати повністю газоподібною.
Водневі зв'язки є важливими силами біополімерів, які включають ДНК, білки та целюлозу. Всі ці полімери втрачають свої рідні структури при кипінні. Водневі зв'язки між амінокислотами, близькими один до одного в первинній структурі, можуть породжувати регулярні повторювані структури, такі як спіралі або складки, в білках (вторинна структура).
Іонні взаємодії
Іонні взаємодії є важливими силами, що стабілізують структуру білка, що виникають внаслідок іонізації R-груп в амінокислотах, що містять білок. До них відносяться карбоксильні амінокислоти (ТУТ), амінокислоти аміну, а також сульфгідрил цистеїну і іноді гідроксил тирозину.
Гідрофобні сили
Гідрофобні сили стабілізують структуру білка в результаті взаємодій, які сприяють виключенню води. Неполярні амінокислоти (зазвичай зустрічаються у внутрішній частині білків) сприяють асоціації один з одним, і це має ефект виключення води. Виключена вода має більш високу ентропію, ніж вода, що взаємодіє з гідрофобними бічними ланцюгами. Це пояснюється тим, що вода вирівнюється дуже регулярно і чітко виражено при взаємодії з гідрофобними молекулами.
Коли воді забороняється мати такі види взаємодій, набагато більш невпорядковано, що було б, якби вона могла асоціюватися з гідрофобними регіонами. Частково з цієї причини гідрофобні амінокислоти знаходяться в білкових інтер'єрах - так вони можуть виключити воду і підвищувати ентропію.
дисульфідні зв'язки
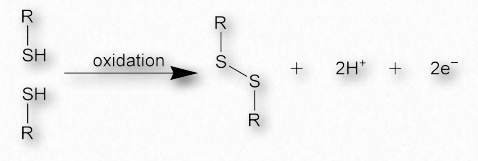
Дисульфідні зв'язки, які виробляються, коли дві сульфгідрильні бічні ланцюги цистеїну наближаються до безпосередньої близькості, ковалентно з'єднуються між собою різні білкові області і можуть надати велику міцність загальній структурі (рис. 2.42 & 2.43). Ода білкової структури Кевіна Ахерна Двадцять років аміно А Визначте білок багатьма способами Їх порядок в пептидному ланцюжку Визначає форми, що білки набирають І коли вони згортаються, це залишає мене веселим Cuz, що робить структури вторинними Це третинний, Мені кажуть, що це відбувається, коли білок складається Але складені ланцюги абсолютно страшні Коли зібрані четвертинні Вони чудеса природи, це точно Створення проблем, виготовлення ліків Дурень може модувати пептидні вірші Але білки походять з рибосом Ці приєднані залишки цистеїну іноді називають цистином. Дисульфідні зв'язки є найсильнішими з сил, що стабілізують білкову структуру.

ван дер Ваальса сили
ван дер Ваальса сили - це термін, який використовується для опису різних слабких взаємодій, у тому числі спричинених притяганням між полярною молекулою та перехідним диполем, або між двома тимчасовими диполями. сили ван дер Ваальса динамічні через коливання характеру тяжіння, і, як правило, слабкі порівняння з ковалентними зв'язками, але може, на дуже коротких відстанях, бути значним.
Посттрансляційні модифікації
Посттрансляційні модифікації можуть призвести до утворення ковалентних зв'язків, що стабілізують білки. Гідроксилювання лізину і проліну в нитках колагену може призвести до зшивання цих груп, а отримані ковалентні зв'язки сприяють зміцненню та стабілізації колагену.
складні моделі
Дві популярні моделі згортання білків в даний час знаходяться під дослідженням. У першому (модель дифузійного зіткнення), подія зародження починає процес, після чого відбувається утворення вторинної структури. Зіткнення між вторинними структурами (як на β-шпильці на рис. 2.37) дозволяють почати складку. На відміну від цього, в моделі нуклеації-конденсації вторинні і третинні структури утворюються разом.
Згортання білків відбувається досить швидко (від 0,1 до 1000 секунд) і може відбуватися під час синтезу - амінокінцевий білок може почати складатися ще до того, як карбоксильний кінець навіть зроблений, хоча це не завжди так.
процес складання
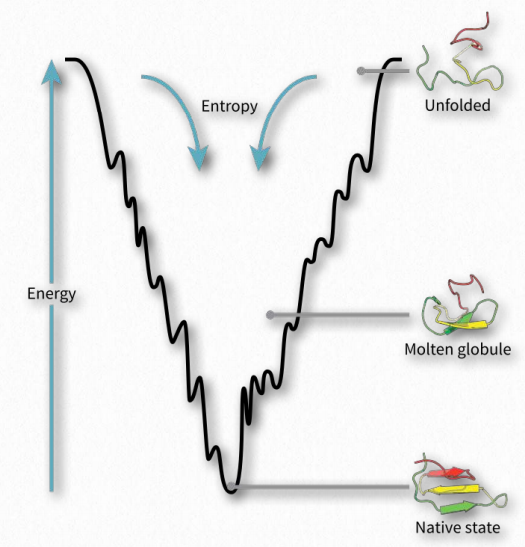
Підтверджується, що згортання білка відбувається в енергетичному ландшафті «складчастої воронки», в якому нативний стан згорнутого білка відповідає мінімальній вільній енергії, можливій в умовах середовища (зазвичай водного розчинника), в якому білок розчиняється. Як видно на діаграмі (рис. 2.44), енергетична воронка має численні локальні мінімуми (провали), в яких складаний білок може потрапити в пастку, коли він рухається вниз по енергетичній ділянці. Інші фактори, такі як температура, електричні/магнітні поля та просторові міркування, ймовірно, відіграють роль.
Якщо зовнішні сили впливають на місцеві мінімуми енергії під час згортання, можна вплинути на процес і кінцевий продукт. Оскільки швидкість автомобіля, що йде по дорозі, вплине на безпеку подорожі, так і енергетичні міркування впливають і направляють процес складання, в результаті чого повністю функціональні, правильно складені білки в деяких випадках і неправильно складені «помилки» в інших.
Застрягання
Оскільки процес згортання протікає до енергетичного мінімуму (дно воронки на рис. 2.44), білок може «застрягти» в будь-якому з локальних мінімумів і не досягти остаточного складеного стану. Хоча складчастий стан, як правило, більш організований і тому має знижену ентропію, ніж розгорнутий стан, є дві сили, які долають зменшення ентропії і рухають процес вперед.
Перший - це величина зменшення енергії, як показано на графіку. Оскільки ΔG = ΔH -TΔS, зменшення ΔH може подолати негативний ΔS, щоб зробити ΔG негативним і проштовхнути процес складання вперед. Сприятливі (знижені) енергетичні умови виникають з утворенням в процесі згортання іонних зв'язків, водневих зв'язків, дисульфідних зв'язків, металевих зв'язків. Крім того, гідрофобний ефект збільшує ентропію, дозволяючи гідрофобним амінокислотам всередині складеного білка виключити воду, тим самим протидіючи впливу впорядкування структури білка, роблячи ΔS менш негативним.
Прогнозування структури
Комп'ютерні програми дуже добре прогнозують вторинну структуру виключно на основі амінокислотної послідовності, але борються з визначенням третинної структури, використовуючи ту ж інформацію. Частково це пов'язано з тим, що вторинні структури мають повторювані точки стабілізації на основі геометрії і будь-яка правильна вторинна структура (наприклад, α-спіраль) дуже мало змінюється від однієї до іншої. Складчасті конструкції, однак, мають величезну кількість можливих структур, як показав Парадокс Левінталя.
Спектроскопія
Через нашу нездатність точно передбачити третинну структуру на основі амінокислотної послідовності, білкові структури фактично визначаються за допомогою методів спектроскопії. У цих підходах білки піддаються різним формам електромагнітного випромінювання, і способи їх взаємодії з випромінюванням дозволяє дослідникам визначати атомні координати при роздільній здатності Ангстрема з щільності електронів (див. Рентгенівська кристалографія) і як ядра спини взаємодіють (див. ЯМР).
парадокс Левінталя
В кінці 1960-х років Сайрус Левінталь окреслив величину складності проблеми згортання білка. Він зазначив, що для білка з 100 амінокислотами він матиме 99 пептидних зв'язків і 198 міркувань для φ і ψ кутів. Якби кожен з них мав лише три конформації, це призвело б до 3198 різних можливих складок або 2,95x1094.
Навіть допускаючи розумний проміжок часу (одна наносекунда) для кожної можливої складки, це займе більше часу, ніж вік Всесвіту, щоб вибірка всіх з них, що чітко означає, що процес згортання не відбувається послідовним випадковим вибіркою і що спроби визначити структуру білка шляхом випадкові вибірки були приречені на провал. Тому Левінталь запропонував, що згортання відбувається послідовним процесом, який починається з події зародження, яка спрямовує процес швидко і не відрізняється від процесу воронки, зображеного на малюнку 2.44.
Захворювання неправильного згортання білка
Правильне складання білків має важливе значення для їх функції. Звідси випливає, що неправильне згортання білків (також називається протеопатією) може мати наслідки. У деяких випадках це може просто призвести до неактивного білка. Неправильне згортання білка також відіграє певну роль у численних захворюваннях, таких як Божевільна коров'яча хвороба, хвороба Альцгеймера, хвороба Паркінсона та хвороба КрейцфельдЯкоба. Багато, але не всі, неправильні захворювання вражають тканини мозку.
нерозчинні відкладення
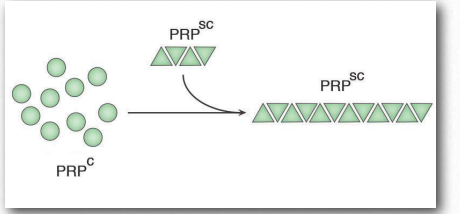
Неправильно складені білки зазвичай утворюють агрегати, звані амілоїдами, які шкідливі для тканин, що містять їх, оскільки вони змінюються від розчинних до нерозчинних у воді та утворюють відкладення. Процес, за допомогою якого відбувається неправильне складання (рис. 2.45), не зовсім зрозумілий, але в багатьох випадках було продемонстровано, що «насіннєвий» білок, який неправильно складається, може викликати таке ж неправильне згортання в інших копіях того ж білка. Ці білки насіння відомі як пріони, і вони діють як інфекційні агенти, що призводить до поширення хвороби. Список захворювань людини, пов'язаних з неправильним складанням білка, довгий і продовжує зростати. Посилання на Вікіпедію тут.
Пріони
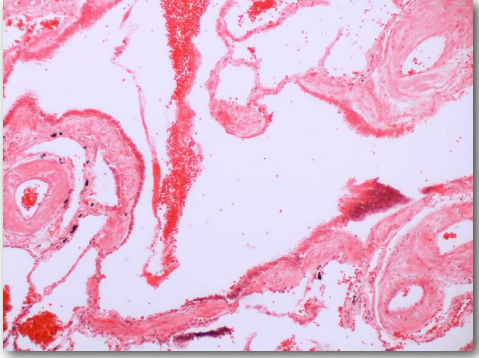
Пріони - це інфекційні білкові частинки, що викликають трансмісивні губчасті енцефалопатії (ТСЕ), найвідомішою з яких є хвороба Божевільної корови. Інші прояви включають захворювання, вискоблювання, у овець та хвороби людини, такі як хвороба КрейтцфельдтЯкоба (CJD), фатальна сімейна безсоння та куру. Білок, який бере участь у цих захворюваннях, є мембранним білком під назвою PrP. PrP кодується в геномі багатьох організмів і знаходиться в більшості клітин організму. PrPC - це назва, дана структурі PrP, яка є нормальною і не пов'язаної з хворобою. PrPSC - це назва, що дається неправильно складеної формі того ж білка, що пов'язано з розвитком симптомів захворювання (рис. 2.45).
неправильно складено
Помилковий PrPSC пов'язаний із захворюваннями TSE і діє як інфекційна частинка. Третя форма PRP, звана PRpREs, може бути виявлена в ЦЕС, але не є інфекційною. «res» PrPres вказує на те, що він стійкий до протеази. Варто відзначити, що всі три форми PrP мають однакову амінокислотну послідовність і відрізняються один від одного тільки способами згортання поліпептидних ланцюгів. Найбільш небезпечною помилковою формою PrP є PrPSc, через свою здатність діяти як інфекційний агент - насіннєвий білок, який може викликати неправильне згортання PrPC, перетворюючи таким чином його в PrPSc.
Функція
Функція PrPC невідома. Миші, які не мають гена PrP, не мають великих відхилень. Вони, здається, виявляють проблеми з довгостроковою пам'яттю, пропонуючи функцію для PRPC. Стенлі Прусінер, який відкрив пріони і придумав термін, отримав Нобелівську премію з медицини в 1997 році за свою роботу. Я думаю, що якби я мав шанс бути на білку, складаючи пріон, я б крутив його і заради добра Зупиніть його робити помилки
Амілоїди
Амілоїди - це сукупність неправильно згорнутих білкових агрегатів, які знаходяться в організмі людини. Як наслідок їх неправильної складаності, вони нерозчинні і сприяють близько двадцяти захворювань людини, включаючи важливі неврологічні ті, що включають пріони. Захворювання включають (уражений білок в дужках) - хвороба Альцгеймера (Амілоїд β), хвороба Паркінсона (α-синуклеїн), хвороба Хантінгтона (хантінгтин), ревматоїдний артрит (сироватковий амілоїд А), смертельне сімейне безсоння (PrPSC) та інші.
Амінокислотна послідовність відіграє певну роль в амілоїдогенезі. Поліпептиди, багаті глутаміном, поширені в дріжджах і людських пріонами. Тринуклеотидні повторення важливі при хворобі Хантінгтона. Там, де послідовність не є фактором, гідрофобна асоціація між β-листами може зіграти певну роль.
Амілоїд β
Амілоїд β відноситься до колекцій невеликих білків (36-43 амінокислот), які, здається, відіграють певну роль у хворобі Альцгеймера. (Тау-білок є іншим фактором.) Насправді вони є основними компонентами амілоїдних бляшок, що знаходяться в мозку пацієнтів, які страждають на хворобу, і виникають внаслідок протеолітичного розщеплення більшого глікопротеїду-попередника амілоїду, який називається протеїн-попередник амілоїду, інтегрального мембранного білка нервових клітин, функція якого невідома. Дві протеази, β-секретаза і γ- секретаза виконують цю функцію. Амілоїдні β білки неправильно згортаються і, здається, спонукають інші білки до неправильного складання і, таким чином, випадають в осад і утворюють амілоїд, характерний для захворювання. Бляшки токсичні для нервових клітин і породжують деменцію, характерну для захворювання.
Вважається, що агрегація білків амілоїду β при неправильному складанні призводить до утворення активних форм кисню, і це засіб, за допомогою якого нейрони пошкоджуються. Невідомо, яка фактична функція амілоїду β. Аутосомно-домінантні мутації в білку призводять до раннього початку захворювання, але це відбувається не більше ніж в 10% випадків. Стратегії лікування захворювання включають інгібування секретаз, які генерують пептидні фрагменти з білка попередника амілоїду.
Хантінгтін
Хантінгтін є центральним геном при хворобі Хантінгтона. Білок, виготовлений з нього, багатий глютаміном, з 6-35 таких залишків у формі дикого типу. При хворобі Хантінгтона цей ген мутується, збільшуючи кількість глютамінів у мутантному білку до між 36 і 250. Розмір білка змінюється в залежності від кількості глютамінів в мутантному білку, але білок дикого типу має понад 3100 амінокислот і молекулярну масу близько 350 000 Да. Його точна функція невідома, але хантінгтин знаходиться в нервових клітині, з найвищим рівнем в головному мозку. Вважається, що він може відігравати роль у транспорті, сигналізації та захисті від апоптозу. Хантінгтін також необхідний для раннього ембріонального розвитку. Всередині клітини виявляється хантінгтін, локалізований переважно з мікротрубочками і везикулами.
Тринуклеотид повторити
Ген хантінгтину містить багато копій послідовності CAG (називається тринуклеотидними повтореннями), які кодують багато глютамінів у білку. Хвороба Хантінгтона виникає, коли при копіюванні ДНК гена генеруються зайві копії послідовності CAG. Розширення повторюваних послідовностей може відбуватися через ковзання полімерази щодо шаблону ДНК при реплікації. В результаті може бути зроблено кілька додаткових копій повтору тринуклеотиду, в результаті чого утворюються білки зі змінною кількістю залишків глютаміну. До 35 повторів можна переносити без проблем. Кількість повторів може розширюватися протягом життя людини, правда, за тим же механізмом. Особи з 36-40 повторами починають проявляти ознаки захворювання і якщо їх більше 40, хвороба буде присутній.
Молекулярні шаперони
Важливість правильного згортання білків підкреслюється захворюваннями, пов'язаними з неправильно згорнутими білками, тому не дивно, що клітини витрачають енергію, щоб полегшити правильне згортання білків. Клітини використовують два класи білків, відомих як молекулярні шаперони, щоб полегшити таке згортання в клітині. Молекулярні шаперони бувають двох видів, шаперони та шапероніни. Прикладом першої категорії є клас білків Hsp70. Hsp розшифровується як «білок теплового шоку», виходячи з того, що ці білки вперше спостерігалися у великій кількості в клітині, які були ненадовго піддані високим температурам. Функція HSPS допомагає клітинам у стресах, що виникають внаслідок теплового шоку та впливу окислювальних умов або токсичних важких металів, таких як кадмій та ртуть. Однак вони також відіграють важливу роль у нормальних умовах, де вони допомагають правильному складанню поліпептидів, запобігаючи аберрантним взаємодіям, які можуть призвести до неправильної згортання або агрегації. Білки Hsp70 знаходяться майже у всіх клітині і використовують гідроліз АТФ для стимуляції структурних змін форми шаперону для розміщення зв'язування білків субстрату. Зв'язуючий домен HSP70s містить β-барельну структуру, яка обертається навколо поліпептидного ланцюга субстрату і має спорідненість до гідрофобних бічних ланцюгів амінокислот. Як показано на малюнку 2.50, Hsp70 зв'язується з поліпептидами, коли вони виходять з рибосом під час синтезу білка. Зв'язування субстрату стимулює гідроліз АТФ, і цьому сприяє інший білок теплового шоку, відомий як Hsp40. Гідроліз АТФ призводить до того, що Hsp70 приймається на закриту конформацію, яка допомагає захистити відкриті гідрофобні залишки та запобігти агрегації або місцевому неправильному складанню.
Після завершення синтезу білка АДФ вивільняється і замінюється АТФ, і це призводить до вивільнення білка субстрату, який потім дозволяє поліпептиду повної довжини складатися правильно.
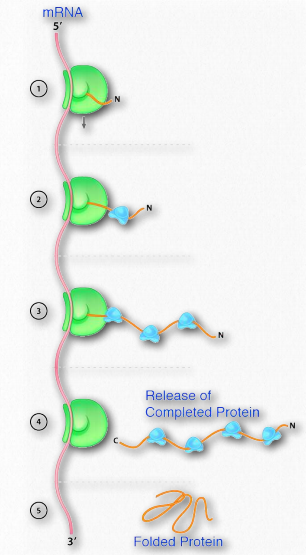
При тепловому ударі
У часи теплового шоку або окислювального стресу білки Hsp70 зв'язуються з розгорнутими гідрофобними ділянками білків, щоб аналогічно запобігти їх агрегації та дозволяючи їм належним чином перегортатися. Коли білки пошкоджуються, Hsp70 набирає ферменти, які вбивають пошкоджений білок, щоб націлити їх для руйнування в протеасомах. Таким чином, білки Hsp70 відіграють важливу роль у забезпеченні не тільки правильного складання білків, але й видалення пошкоджених або нефункціональних білків шляхом деградації в протеасомі.
Шапероніни
Другий клас білків, які беруть участь у наданні допомоги іншим білкам, щоб скласти належним чином, відомі як шапероніни. Існує дві первинні категорії шаперонінів - I клас (міститься в бактеріях, хлоропластах і мітохондріях) і II клас (міститься в цитозолі еукаріотів і архебактерій). Найбільш вивченими шаперонінами є складні білки Groel/Groes, виявлені в бактеріях (рис. 2.51).
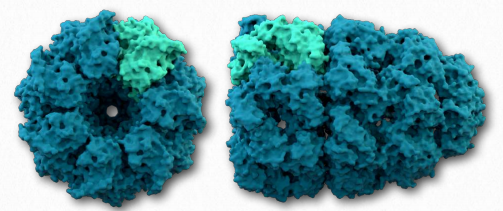
Groel/Groes може не бути в змозі скасувати агреговані білки, але, полегшуючи правильне складання, він забезпечує конкуренцію за неправильне складання як процес і може зменшити або усунути проблеми, що виникають внаслідок неправильного складання. GroEl - це подвійне кільце 14mer з гідрофобною областю, яка може полегшити складання субстратів розміром 15-60 кДа. GroEs - це одиночний гептамер, який зв'язується з GroEL при наявності АТФ і функціонує як обкладинка над GroEL. Гідроліз АТФ шаперонінами викликає великі конформаційні зміни, які впливають на зв'язування білків субстрату і їх згортання. Точно невідомо, як шапероніни складають білки. Пасивні моделі постулюють комплекс шапероніну, що функціонує інертно, запобігаючи несприятливим міжмолекулярним взаємодіям або встановлюючи обмеження на простори, доступні для складання. Активні моделі дозволяють припустити, що структурні зміни в комплексі шапероніну викликають структурні зміни білка субстрату.
Розпад білка
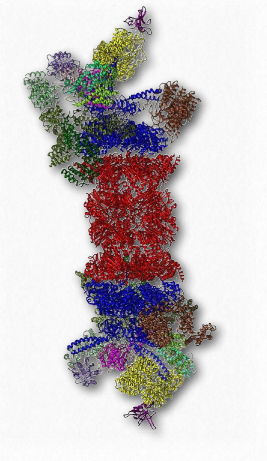
Ще одним білковим комплексом, який виконує важливу функцію в життєвій динаміці білків, є протеасома (рис. 2.52). Протеасоми, які є у всіх еукаріотів і археанів, а також деякі бактерії, функціонують для руйнування непотрібних або пошкоджених білків шляхом протеолітичної деградації. Протеасоми допомагають регулювати концентрацію деяких білків і деградують ті, які неправильно складаються. Шляхи протеазомальної деградації відіграє важливу роль у клітинних процесах, які включають прогресування через клітинний цикл, модуляцію експресії генів та відповідь на окислювальні стреси.
Деградація в протеасомі дає короткі пептиди довжиною від семи до восьми амінокислот. Треонінові протеази грають важливу роль. Розщеплення цих пептидів дає окремі амінокислоти, тим самим полегшуючи їх переробку в клітині. Білки орієнтовані на деградацію в еукаріотичних протеасомах шляхом приєднання до множинних копій невеликого білка під назвою убиквітин (8,5 кДа - 76 амінокислот). Фермент, що каталізує реакцію, відомий як убіквітин лігаза. Отримана ланцюг поліубіквітину пов'язана протеасомою і починається деградація. Убіквітин був названий завдяки тому, що він повсюдно зустрічається в еукаріотичних клітині.
Убіквітин
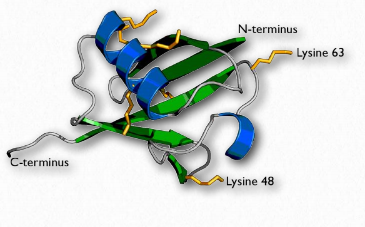
Убіквітин (рис. 2.53) - це невеликий (8,5 кДа) багатофункціональний білок, який міститься в клітинах-еукаріотів. Зазвичай його додають до цільових білків під дією ферментів убиквітинлігази (Е3 на рис. 2.54). Може бути додана одна (убіквітинація) або багато (поліубиквітинація) молекул убіквітину. Приєднання убіквітину відбувається через бічний ланцюг одного з семи різних залишків лізину в убиквітині.
Додавання убіквітину до білків має багато ефектів, найвідомішим з яких є націлювання білка на деградацію протеасоми. Протеасомальний таргетинг спостерігається, коли поліубиквітинація відбувається при лізини #29 і 48. Полюбіквітинація або монобіквітинація при інших лізини може призвести до зміни клітинного розташування та зміни білково-білкових взаємодій. Останнє може змінити вплив на запалення, ендоцитарний обіг, трансляцію та відновлення ДНК.
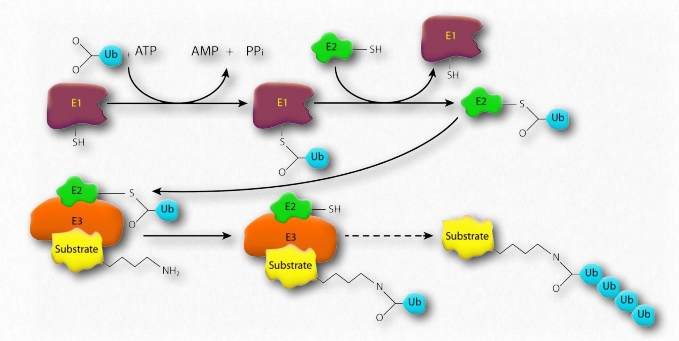
Несправність лігази Убіквітин
Паркін - це білок, пов'язаний із хворобою Паркінсона, який при мутації пов'язаний із спадковою формою захворювання, що називається аутосомно-рецесивною хворобою ювенільного Паркінсона. Функція білка невідома, але він є компонентом системи лігази E3 ubiquitin ligaze, що відповідає за перенесення убиквітину з білка Е2 в бічний ланцюг лізину на бік-мішені. Вважається, що мутації в паркіні призводять до протеазомальної дисфункції і, як наслідок, нездатності розщеплювати білки, шкідливі для дофамінергічних нейронів. Це призводить до загибелі або збою цих нейронів, що призводить до хвороби Паркінсона.
Іскрометно невпорядковані білки
Фільм 2.1 - Динамічний рух цитохрому С у розчині Вікіпедії
Як видно з численних прикладів, описаних в інших місцях книги, 3-D структура білків важлива для їх функції. Але, все частіше, стає очевидним, що не всі білки згортаються в стійку структуру. Дослідження так званих іскромно невпорядкованих білків (ВПО) протягом останніх десятиліть показали, що багато білків біологічно активні, навіть вважаючи, що вони не згортаються в стабільні структури. Проте інші білки демонструють регіони, які залишаються розгорнутими (області ВПО), навіть коли решта поліпептиду складається в структуровану форму.
По суті, невпорядковані білки та невпорядковані області в білках насправді були відомі протягом багатьох років, але розглядалися як аномалія. Лише нещодавно, усвідомлюючи, що регіони ВПО та ВПО широко поширені серед еукаріотичних білків, було визнано, що спостережуваний розлад є «особливістю, а не помилкою».
Фільм 2.2 СУМО-1, білок з іскромно невпорядкованими розділами Вікіпедії
Порівняння ВПО показує, що вони поділяють характеристики послідовності, які, здається, сприяють їх невпорядкованому стану. Тобто, так само, як деякі послідовності амінокислот можуть сприяти згортанню поліпептиду в певну структуру, амінокислотні послідовності ВПО сприяють їх залишкам розгорнутих. Видно, що регіони ВПО мають низький вміст гідрофобних залишків і надзвичайно багаті полярними залишками та проліном. Наявність великої кількості заряджених амінокислот у ВПО може гальмувати згортання за допомогою відштовхування заряду, в той час як відсутність гідрофобних залишків ускладнює формування стійкого гідрофобного ядра, а пролін перешкоджає утворенню спіральних структур. Спостережувані відмінності між послідовностями амінокислот у ВПО та структурованими білками були використані для розробки алгоритмів прогнозування того, чи буде порушена задана послідовність амінокислот.
Яке значення іскромно невпорядкованих білків або регіонів? Той факт, що ця властивість кодується в їх амінокислотних послідовностях, говорить про те, що їх розлад може бути пов'язаний з їх функцією. Гнучкий, рухливий характер деяких регіонів ВПО може відігравати вирішальну роль у їх функції, дозволяючи перехід до складчастої структури при зв'язуванні білкового партнера або зазнає посттрансляційної модифікації. Дослідження кількох відомих білків з регіонами ВПО пропонують деякі відповіді. Регіони ВПО можуть підвищувати здатність білків, таких як лак-репресор, транслокуватися вздовж ДНК для пошуку конкретних місць зв'язування. Гнучкість ВПО також може бути активом у білково-білкових взаємодіях, особливо для білків, які, як відомо, взаємодіють з багатьма різними білковими партнерами.
Наприклад, p53 має області ВПО, які можуть дозволити білку взаємодіяти з різними функціональними партнерами. Порівняння відомих функцій білків з прогнозами розладу в цих білках свідчить про те, що ВПО та ВПО можуть непропорційно функціонувати в сигналізації та регуляції, тоді як більш структуровані білки перекошуються в бік ролі в каталізі та транспорті. Цікаво, що багато білків, виявлених як в рибосомах, так і в спліцеосомах, передбачається, що мають ВПО області, які можуть відігравати роль у правильній збірці цих комплексів. Незважаючи на те, що ВПО не дуже довго вивчалися інтенсивно, те, що про них мало що відомо, свідчить про те, що вони відіграють важливу і недооцінену роль у клітині.
метаморфічні білки
Ще однією групою білків, які останнім часом змінили наше уявлення про структуру і функції білка, є так звані метаморфічні білки. Ці білки здатні утворювати більше одного стабільного, складеного стану, починаючи з однієї амінокислотної послідовності. Хоча це правда, що множинні складчасті конформації не виключені законами фізики та хімії, метаморфічні білки є відносно новим відкриттям. Звичайно, було відомо, що білки пріонів здатні складатися в альтернативні структури, але метаморфічні білки, здається, здатні перемикатися вперед і назад між двома стабільними структурами. Хоча в деяких випадках метаморфічний білок зазнає цього перемикання у відповідь на зв'язування іншої молекули, деяких білків, які можуть здійснити цей перехід самостійно. Цікавим прикладом є сигнальна молекула, лімфотактин. Лімфотактин має дві біологічні функції, які здійснюються двома його конформерами - мономерна форма, яка зв'язує рецептор лімфотактину, і димерну форму, яка зв'язує гепарин. Цілком можливо, що такий вид перемикання більш поширений, ніж вважалося.
Розгортання денатурованих білків
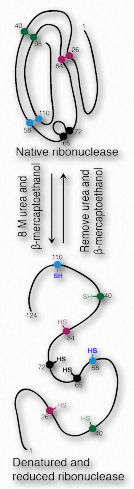
Вся інформація для згортання білка міститься в амінокислотної послідовності білка. Тоді може здатися цікавим, що більшість білків не згортаються у свою належну, повністю активну форму після того, як вони були+++ денатурації та видалення денатуратора. Деякі роблять, насправді. Одним з наочних прикладів є рибонуклеаза великої рогатої худоби (рис. 2.55). Його каталітична активність дуже стійка до нагрівання та сечовини, і спроби денатурувати його не дуже добре. Однак, якщо обробляти фермент β-меркаптоетанолом (який розриває дисульфідні зв'язки) перед обробкою сечовиною та/або нагріванням, активність втрачається, що свідчить про те, що ковалентні дисульфідні зв'язки допомагають стабілізувати загальну ферментну структуру, і коли вони порушені, денатурація може легко відбутися. Коли суміш охолоне назад до кімнатної температури, з часом деяка активність ферментів знову з'являється, що свідчить про те, що рибонуклеаза знову згортається в нових умовах.
Цікаво, що ренатурація відбудеться максимально, якщо під час процесу в розчині залишиться крихітна кількість β-меркаптоетанолу. Причина цього полягає в тому, що β- меркаптоетанол дозволяє зменшити (і розірвати) випадкових, неправильних дисульфідних зв'язків під час процесу згортання. Без нього ці дисульфідні зв'язки будуть перешкоджати утворенню належних складок.
необоротна денатурація
Більшість ферментів, однак, не поводяться як рибонуклеаза великої рогатої худоби. Після денатурації їх активність не може бути відновлена до будь-якого значного Існує не дуже багато способів Інактивація РНКази Це стабільно, коли жарко або холодно Оскільки дисульфіди міцно тримаються Якщо ви хочете змусити його затримати Використовуйте гарячий ступінь меркаптоетанолу. Це може здатися суперечити ідеї згортання інформації, притаманної послідовності амінокислот в білку. Це не так.
Більшість ферментів не відновлюються належним чином після денатурації з двох причин. По-перше, нормальне згортання може відбуватися, коли виробляються білки. Взаємодії між амінокислотами на початку синтезу не «плутаються» взаємодіями з амінокислотами пізніше в синтезі, оскільки ці амінокислоти не присутні, як процес починається.
роль шапероніна
В інших випадках процес згортання деяких білків в клітці спирався на дію білків шапероніну (див. ТУТ). За відсутності шаперонінів відбуваються взаємодії, які можуть призвести до неправильної складки, тим самим запобігаючи правильному складанню. Таким чином, раннє складання та допомога шаперонінів усувають деякі потенційні взаємодії «неправильної складаності», які можуть виникнути, якщо вся послідовність була присутня, коли почалося складання.
четвертинна структура
Четвертий рівень білкової структури - це четвертинна структура. Він відноситься до структур, які виникають в результаті взаємодії між множинними поліпептидами. Одиниці можуть бути ідентичними множинними копіями або можуть являти собою різні поліпептидні ланцюги. Гемоглобін для дорослих є хорошим прикладом білка з четвертинною структурою, який складається з двох однакових ланцюгів, що називаються α, і двох однакових ланцюгів, що називаються β.
Хоча α-ланцюги дуже схожі на β- ланцюги, вони не ідентичні. Обидва α- і β-ланцюга також пов'язані з єдиним поліпептидним ланцюгом у спорідненому білку під назвою міоглобін. І міоглобін, і гемоглобін мають схожість в зв'язуванні кисню, але їх поведінка по відношенню до молекули значно відрізняється Примітно, що множинні субодиниці гемоглобіну (з четвертинною структурою) порівняно з єдиною субодиницею міоглобіну (без четвертинної структури) призводять до цих відмінностей.
Посилання
1. https://en.wikipedia.org/wiki/Van_der_W аалс_форс 105