2.2: Структура та функції - Амінокислоти
- Page ID
- 2492

Джерело: BiochemFFA_2_1.pdf. Весь підручник доступний безкоштовно від авторів за адресою http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
Всі білки на лицьовій поверхні землі складаються з тих же 20 амінокислот. Пов'язані між собою в довгі ланцюги, які називаються поліпептидами, амінокислоти є будівельними блоками для великого асортименту білків, знайдених у всіх живих клітині.

«Це одне з найяскравіших узагальнень біохімії... що двадцять амінокислот та чотири основи, з незначними застереженнями, однакові у всій Природі». - Френсіс Крик
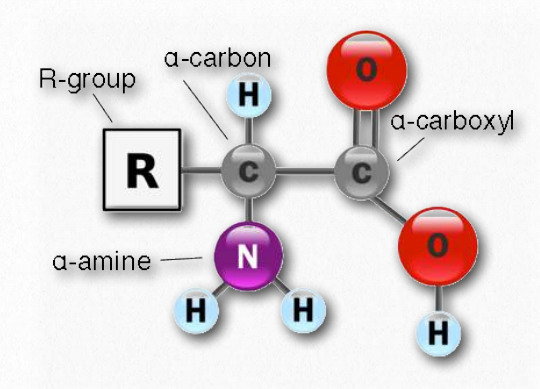
Всі амінокислоти мають однакову основну будову, яка показана на малюнку 2.1. У «центрі» кожної амінокислоти знаходиться вуглець, званий α вуглецем і приєднані до нього чотири групи - водень, α- карбоксильна група, α-амінна група і R-група, яку іноді називають бічним ланцюгом. Вуглецеві, карбоксильні та аміногрупи є загальними для всіх амінокислот, тому R-група є єдиною унікальною особливістю кожної амінокислоти. (Незначним винятком з цієї структури є пролін, в якому кінець R-групи приєднаний до α-аміну.) За винятком гліцину, який має R-групу, що складається з атома водню, всі амінокислоти в білках мають чотири різні групи, прикріплені до них і, отже, можуть існувати в двох дзеркальних формах відображення, L і D. За лише дуже незначними винятками, кожна амінокислота, що міститься в клітині і в білках, знаходиться в конфігурація L.
Є 22 амінокислоти, які містяться в білках, і з них тільки 20 вказані універсальним генетичним кодом. Інші, селеноцистеїн та піролізин, використовують тРНК, які здатні базувати пару зі стоп-кодонами в мРНК під час трансляції. Коли це станеться, ці незвичайні амінокислоти можуть бути включені в білки. Ферменти, що містять селеноцистеїн, наприклад, включають глутатіонпероксидази, тетрайодтироніну 5' дейодинази, тіоредоксиредуктази, форміатдегідрогенази, гліцин-редуктази та селенофосфатсинтетазу. Білки, що містять піролізин, зустрічаються набагато рідше і здебільшого приурочені до архей.
Ефірні та несуттєві
Дієтологи поділяють амінокислоти на дві групи - незамінні амінокислоти (повинні бути в раціоні, оскільки клітини не можуть їх синтезувати) і незамінні амінокислоти (можуть бути зроблені клітинами). Ця класифікація амінокислот має мало спільного зі структурою амінокислот. Незамінні амінокислоти значно варіюються від одного організму до іншого і навіть відрізняються у людини в залежності від того, дорослі вони або діти. У таблиці 2.1 наведені незамінні і незамінні амінокислоти у людини.
Деякі амінокислоти, які зазвичай не є незамінними, може знадобитися отримувати з раціону в певних випадках. Особи, які не синтезують достатню кількість аргініну, цистеїну, глютаміну, проліну, селеноцистеїну, серину та тирозину, наприклад, можуть потребувати харчових добавок, що містять ці амінокислоти.
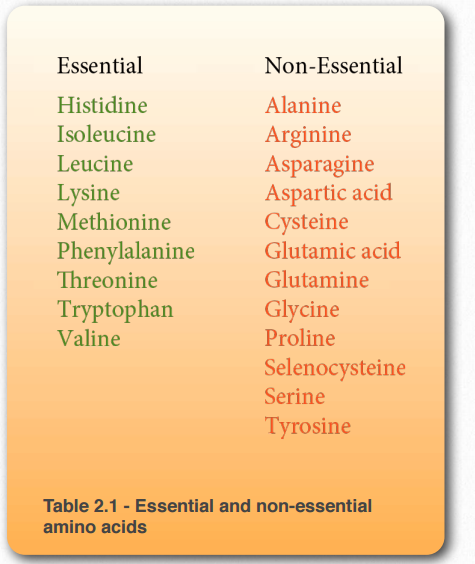
Таблиця 2.1 - Незамінні і незамінні амінокислоти
Небілкові амінокислоти
Існують також α-амінокислоти, знайдені в клітині, які не включені в білки. До поширених відносяться орнітин і цитрулін. Обидва ці сполуки є проміжними продуктами в циклі сечовини. Орнітин є метаболічним попередником аргініну, а цитрулін може вироблятися при розпаді аргініну. Остання реакція виробляє оксид азоту, важливу сигнальну молекулу. Цитрулін є побічним продуктом метаболізму. Іноді його використовують як харчову добавку для зменшення м'язової втоми.
Хімія R-групи
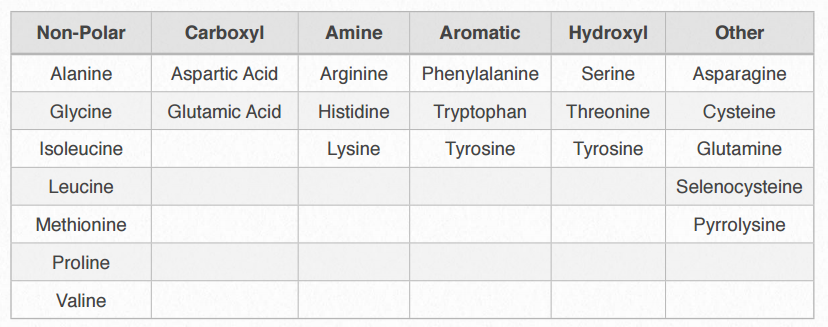
Таблиця 2.2 - Категорії амінокислот (виходячи з властивостей R-групи)
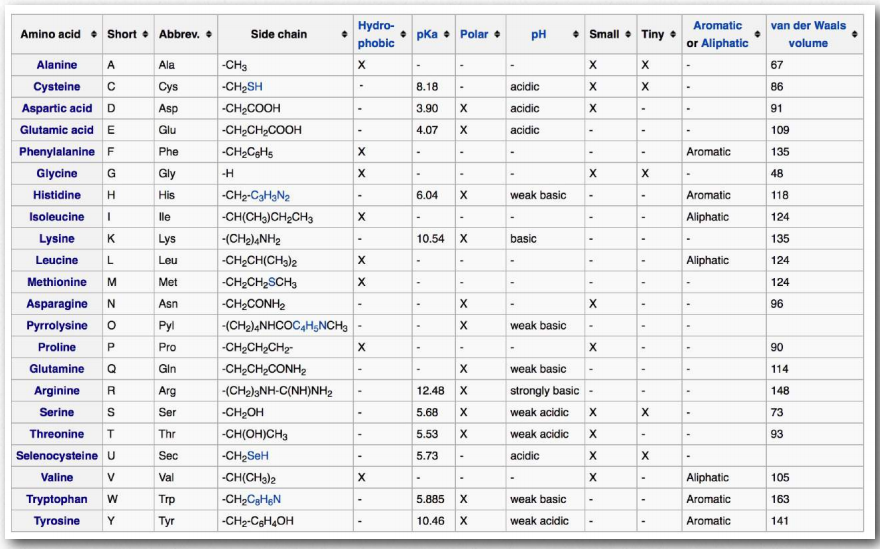
Ми поділяємо амінокислоти на категорії, виходячи з хімії їх R-груп. Якщо порівнювати групи амінокислот у різних підручниках, ви побачите різні назви категорій і (іноді) однакові амінокислоти класифікуються по-різному різними авторами. Дійсно, ми класифікуємо тирозин як ароматичну амінокислоту, так і як гідроксильну амінокислоту. Корисно класифікувати амінокислоти на основі їх R-груп, адже саме ці бічні ланцюги надають кожній амінокислоті свої характерні властивості. Таким чином, амінокислоти з (хімічно) схожими бічними групами можна очікувати функціонування подібним чином, наприклад, під час згортання білка.
неполярні амінокислоти
- Аланін (Ala/A) - одна з найпоширеніших амінокислот, що містяться в білках, посідаючи друге місце лише лейцину за наявністю. D-форма амінокислоти також міститься в клітинних стінках бактерій. Аланін є несуттєвим, легко синтезується з пірувату. Він кодується для GCU, GCC, GCA та GCG.
- Гліцин (Gly/G) - амінокислота з найкоротшим бічним ланцюгом, що має R-групу, що відповідає лише одному водню. В результаті гліцин - єдина амінокислота, яка не є хіральною. Його невеликий бічний ланцюг дозволяє йому легко вписуватися як в гідрофобні, так і в гідрофільні середовища.
- Гліцин вказується в генетичному коді GGU, GGC, GGA і GGG. Це несуттєво для людини.
- Ізолейцин (Ile/I) є незамінною амінокислотою, кодується AUU, AUC, і AUA. Він має гідрофобний бічний ланцюг, а також є хіральним у своєму бічному ланцюгу.
- Лейцин (Leu/L) - це амінокислота з розгалуженим ланцюгом, яка є гідрофобною і незамінною. Лейцин є єдиною дієтичною амінокислотою, яка, як повідомляється, безпосередньо стимулює синтез білка в м'язах, але обережність в порядку, оскільки 1) існують суперечливі дослідження і 2) токсичність лейцину небезпечна, в результаті чого «чотири D»: діарея, дерматит, деменція і смерть. Лейцин кодується шістьма кодонами: UUA, UUG, CUU, CUC, CUA, CUG.
- Метіонін (Мет/М) - незамінна амінокислота, яка є однією з двох сірковмісних амінокислот - цистеїн інший. Метіонін неполярний і кодується виключно кодоном AUG. Це «ініціатор» амінокислоти в синтезі білка, будучи першою включеною в білкові ланцюги. У прокаріотичних клітині формується перший метіонін в білку.
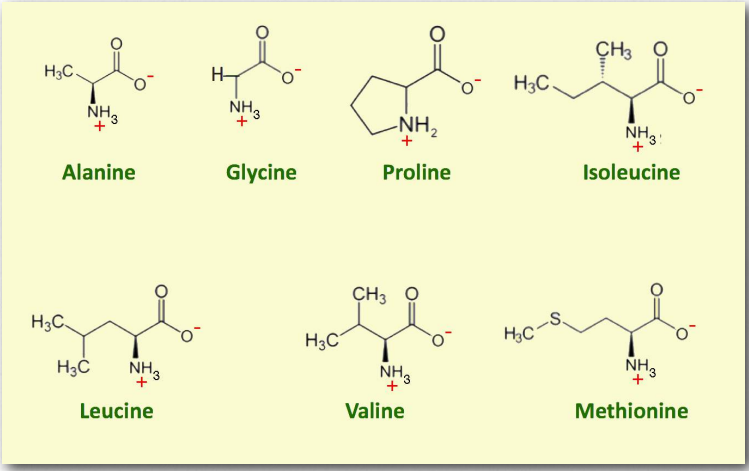
- Пролін (Pro/P) - єдина амінокислота, що міститься в білках з R-групою, яка з'єднується з власною α-аміногрупою, утворюючи вторинний амін і кільце. Пролін є незамінною амінокислотою і кодується CCU, CCC, CCA та CCG. Це найменш гнучкий з білкових амінокислот і, таким чином, дає конформаційну жорсткість, коли він присутній у білку. Наявність проліну в білку впливає на його вторинну структуру. Це руйнівник α-спіралей і β-ниток. Пролін часто гідроксилюється в колагені (реакція вимагає вітаміну С - аскорбат), і це має ефект підвищення конформаційної стабільності білка. Пролін гідроксилювання фактора, індукованого гіпоксією (HIF) служить датчиком рівня кисню і мішенню HIF для руйнування, коли кисню багато.
- Валін (Val/V) - незамінна неполярна амінокислота, що синтезується в рослині. Це примітно в гемоглобіні, оскільки коли він замінює глутамінову кислоту в положенні номер шість, це змушує гемоглобін аномально агрегуватися в умовах низького рівня кисню, що призводить до серповидноклітинної хвороби. Валін кодується в генетичному коді GUU, GUC, GUA і GUG.
Карбоксильні амінокислоти
- Аспарагінова кислота (Asp/D) - незамінна амінокислота з карбоксильною групою в своїй Rгрупі. Він легко виробляється шляхом трансамінації оксалоацетату. При рКа 3,9 бічний ланцюг аспарагінової кислоти негативно заряджається при фізіологічному рН. Аспарагінова кислота визначається в генетичному коді кодонами GAU і GAC.
- Глутамінова кислота (Glu/E), яка кодується GAA та GAG, є незамінною амінокислотою, легко виготовленої шляхом трансамінації α- кетоглутарату. Це нейромедіатор і має R-групу з карбоксильною групою, яка легко іонізується (pKA = 4,1) при фізіологічному рН.

Амінні амінокислоти
- Аргінін (Arg/R) - амінокислота, яка в деяких випадках є незамінною, але несуттєвою в інших. Недоношені діти не можуть синтезувати аргінін. Крім того, хірургічні травми, сепсис та опіки підвищують попит на аргінін. Більшість людей, однак, не потребують добавок аргініну. Бічний ланцюг аргініну містить складну гуанідинієву групу з pKA понад 12, що робить його позитивно зарядженим при клітинному рН. Він кодується шістьма кодонами - CGU, CGC, CGA, CGG, AGA та AGG.
- Гістидин (His/H) є єдиною з білкових амінокислот, що містять функціональну групу імідазолу. Це незамінна амінокислота у людини та інших ссавців. З бічним ланцюгом pKA 6 він може легко змінити свій заряд за допомогою невеликої зміни рН. Протонування кільця призводить до утворення двох структур NH, які можуть бути намальовані як дві однаково важливі резонансні структури.
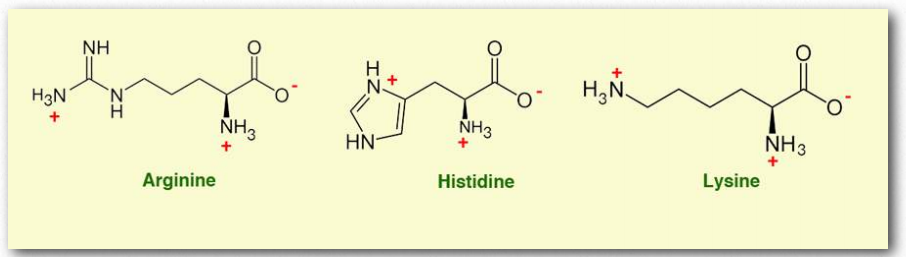
- Лізин (Lys/K) - незамінна амінокислота, кодована AAA та AAG. Він має Rгрупу, яка може легко іонізуватися із зарядом +1 при фізіологічному рН і може бути посттрансляційно модифікована з утворенням ацетиллізину, гідроксилізину та метиллізину. Він також може бути всюдисущим, сумольованим, неддильованим, біотинільованим, карбоксильованим і пупільованим, і. О-глікозилювання гідроксилізину використовується для позначення білків для експорту з клітини. Лізин часто додають в корми для тварин, оскільки він є обмежуючою амінокислотою і необхідний для оптимізації росту свиней і курей.
Ароматичні амінокислоти
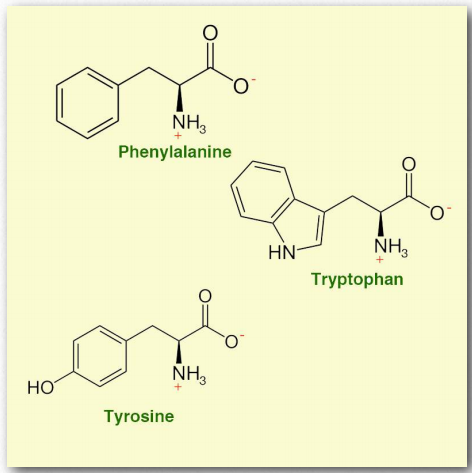
- Фенілаланін (Phe/F) - неполярна незамінна амінокислота, кодована UUU та UUC. Є метаболічним попередником тирозину. Нездатність метаболізувати фенілаланін виникає внаслідок генетичного захворювання, відомого як фенілкетонурія. Фенілаланін є компонентом штучного підсолоджувача аспартаму.
- Триптофан (Trp/W) - незамінна амінокислота, що містить функціональну групу індолу. Це метаболічний попередник серотоніну, ніацину та (у рослин) фітогормону ауксину. Незважаючи на те, що він служить засобом сну, немає чітких результатів досліджень, які вказують на це.
- Тирозин (Tyr/Y) є незамінною амінокислотою, кодованої UAC і UAU. Він є мішенню для фосфорилювання в білках тирозин-протеїнкіназами і відіграє роль у сигнальних процесах. У дофамінергічних клітирах мозку тирозингідроксилаза перетворює тирозин в l-допу, безпосередній попередник дофаміну. Дофамін, в свою чергу, є попередником норадреналіну і адреналіну. Тирозин також є попередником гормонів щитовидної залози і меланіну.
Гідроксильні амінокислоти
- Серин (Ser/S) - одна з трьох амінокислот, що мають R-групу з гідроксилом в ній (треонін і тирозин - інші). Він кодується УКУ, УКС, УЦА, УГК, AGU та AGC. Будучи здатним зв'язуватися з водою водню, він класифікується як полярна амінокислота. Це не важливо для людини. Серин є попередником багатьох важливих клітинних сполук, включаючи пуринів, піримідинів, сфінголіпідів, фолатів та амінокислот гліцину, цистеїну та триптофану. Гідроксильна група серину в білках є мішенню для фосфорилювання певними протеїнкіназами. Серин також входить до складу каталітичної тріади серин-протеаз.
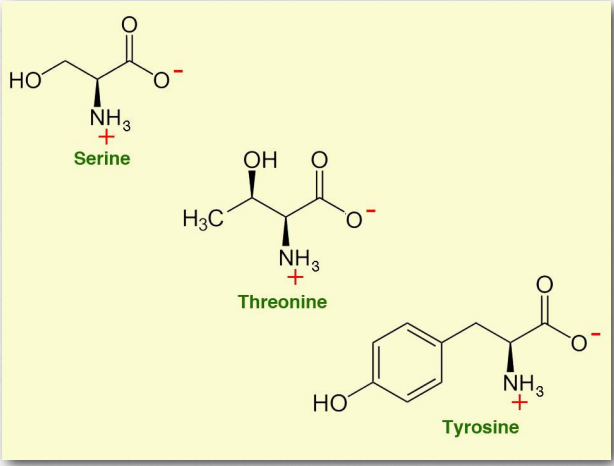
- Треонін (Thr/T) - це полярна амінокислота, яка необхідна. Це одна з трьох амінокислот, що несуть гідроксильну групу (серин і тирозин є іншими) і, як така, є мішенню для фосфорилювання в білках. Він також є мішенню для глікозилювання білків. Треонінові протеази використовують гідроксильну групу амінокислоти в їх каталізі, і вона є попередником в одному біосинтетичному шляху для виготовлення гліцину. У деяких додатках він використовується як про-препарат для підвищення рівня гліцину мозку. Треонін кодується в генетичному коді ACU, ACC, ACA і ACG.
Тирозин - дивіться ТУТ.
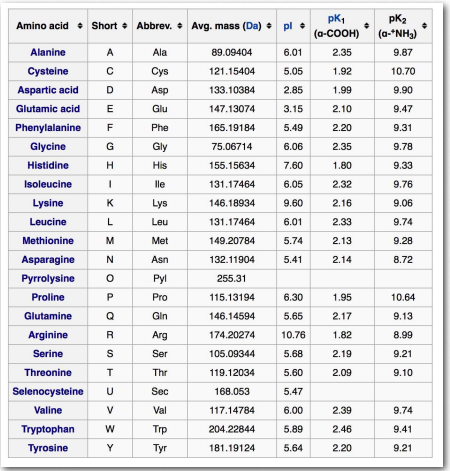
Інші амінокислоти
- Аспарагін (Asn/N) є незамінною амінокислотою, кодованої AAU і AAC. Його карбоксимід в R-групі надає йому полярності. Аспарагін бере участь в утворенні акриламіду в продуктах, приготованих при високих температурах (фритюрі), коли він реагує з карбонільними групами. Аспарагін може бути зроблений в організмі з аспартату шляхом реакції амідації з аміном з глютаміну. Розпад аспарагіну виробляє малат, який може окислюватися в циклі лимонної кислоти.
- Цистеїн (Cys/C) - єдина амінокислота з сульфгідрильною групою в бічному ланцюзі. Це несуттєво для більшості людей, але може бути важливим для немовлят, людей похилого віку та осіб, які страждають певними захворюваннями обміну речовин. Сульфгідрильна група цистеїну легко окислюється до дисульфіду при взаємодії з іншою. Окрім того, що міститься в білках, цистеїн також є компонентом трипептиду, глутатіону. Цистеїн уточнюється кодонами UGU і UGC.
- Глютамин (Gln/Q) - амінокислота, яка зазвичай не є необхідною для людини, але може бути у осіб, які проходять інтенсивні спортивні тренування або з шлунково-кишковими розладами. Він має боковий ланцюг карбоксиміду, який нормально не іонізується під фізіологічними рН, але надає полярність бічному ланцюгу. Глютамин кодується CAA і CAG і легко виробляється шляхом амідації глутамату. Глютамин є найпоширенішою амінокислотою в циркулюючої крові і є однією з небагатьох амінокислот, які можуть перетнути гематоенцефалічний бар'єр.
- Селеноцистеїн (Sec/U) - компонент селенопротеїнів, знайдених у всіх царствах життя. Він є компонентом декількох ферментів, включаючи глутатіонпероксидази і тіоредоксиредуктази. Селеноцистеїн включений в білки за незвичайною схемою, що включає стоп-кодон UGA. Клітини, вирощені за відсутності селену, припиняють синтез білка на УГА. Однак, коли присутній селен, деякі мРНК, які містять послідовність введення селеноцистеїну (SECIS), вставляють селеноцистеїн, коли зустрічається UGA. Елемент SECIS має характерні нуклеотидні послідовності та схеми сполучення основи вторинної структури. Двадцять п'ять людських білків містять селеноцистеїн.
- Піролізин (Pyl/O) - двадцять друга амінокислота, але рідко зустрічається в білках. Як і селеноцистеїн, він не кодується в генетичному коді і повинен бути включений незвичайними засобами. Це відбувається на стоп-кодони UAG. Піролізин міститься в метаногенних архейських організмах і принаймні в одній бактерії, що продукує метан. Піролізин є компонентом ферментів, що продукують метану.
іонізуючі групи
Значення pKA для бічних ланцюгів амінокислот дуже залежать від хімічного середовища, в якому вони присутні. Наприклад, карбоксил R-групи, що міститься в аспарагінової кислоти, має значення pKA 3,9, коли він вільний у розчині, але може досягати 14, коли в певних середовищах всередині білків, хоча це незвично і екстремально. Кожна амінокислота має щонайменше одну іонізуючу амінну групу (α- амін) і одну іонізовану карбоксильну групу (α- карбоксил). Коли вони пов'язані в пептидному зв'язку, вони більше не іонізуються. Деякі, але не всі амінокислоти мають R-групи, здатні іонізуватися. Заряд білка тоді виникає із зарядів α-амінної групи, α- карбоксильної групи. і суми зарядів іонізованих R-груп. Титрування/іонізація аспарагінової кислоти зображено на малюнку 2.10. Іонізація (або деіонізація) в структурі білка може мати значний вплив на загальну конформацію білка і, оскільки структура пов'язана з функцією, великий вплив на активність білка.
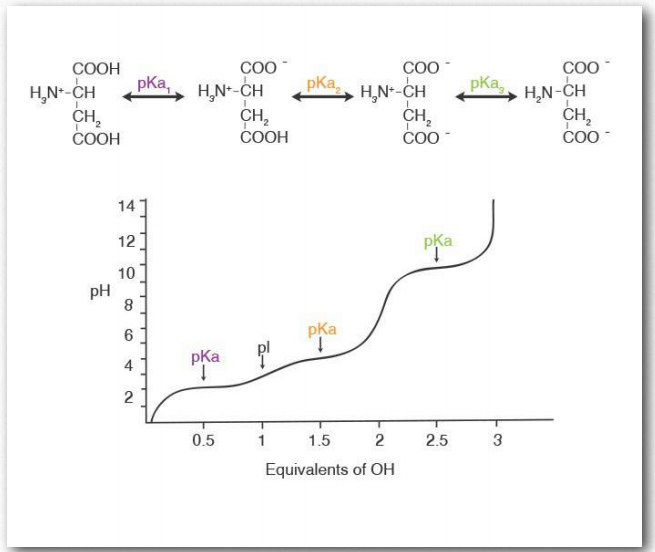
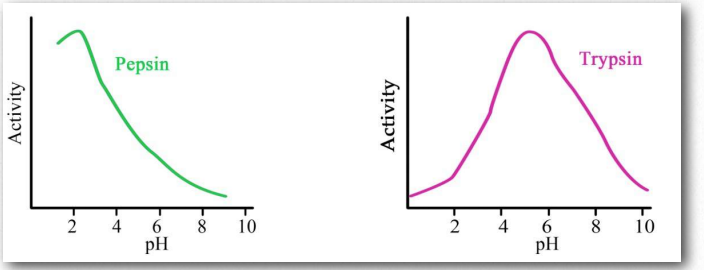
Більшість білків мають відносно вузькі діапазони оптимальної активності, які зазвичай відповідають середовищам, в яких вони знаходяться (рис. 2.11). Варто відзначити, що утворення пептидних зв'язків між амінокислотами видаляє іонізуючі водні як з α- амінної, так і α- карбоксильної груп амінокислот. Таким чином, іонізація/деіонізація в білку виникає тільки з 1) амінокінцевих; 2) карбоксильних кінцевих; 3) R-груп; або 4) інших функціональних груп (таких як сульфати або фосфати), доданих до амінокислот посттрансляційно - див. Нижче.
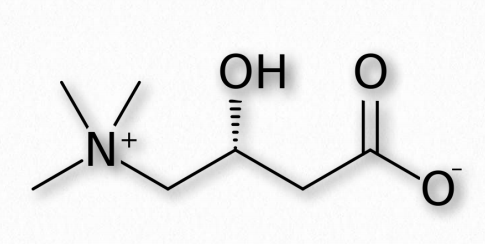
Карнітин
Не всі амінокислоти в клітині містяться в білках. Найбільш поширені приклади включають орнітин (метаболізм аргініну), цитрулін (цикл сечовини) та карнітин (рис. 2.12). Коли жирні кислоти, призначені для окислення, переміщуються в мітохондріон для цієї мети, вони подорожують через внутрішню мембрану, прикріплену до карнітину. З двох стереоізомерних форм активною є L-форма. Молекула синтезується в печінці з лізину і метіоніну.
З екзогенних джерел жирні кислоти повинні бути активовані при попаданні в цитоплазму шляхом приєднання до коферменту А. КоА-частина молекули замінюється карнітином в міжмембранному просторі мітохондріона в реакції, каталізованої карнітинацилтрансферазою I. переноситься через внутрішню мітохондріальну мембрану транслоказом карнітінеацилкарнітину, а потім в матриксі мітохондріона карнітинацилтрансфераза II замінює карнітин коензимом А (рис. 6.88).
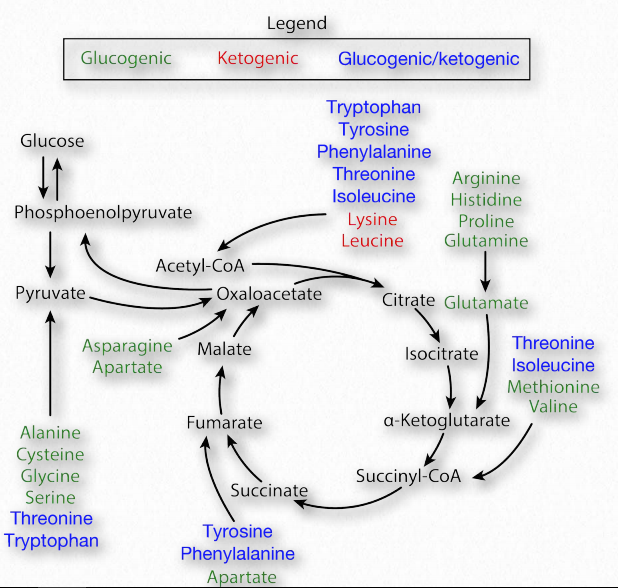
катаболізм амінокислот
Ми класифікуємо амінокислоти як незамінні або несуттєві залежно від того, чи може організм їх синтезувати. Всі амінокислоти, однак, можуть бути розщеплені всіма організмами. Насправді вони є джерелом енергії для клітин, особливо під час голодування або для людей на дієтах, що містять дуже низьку кількість вуглеводів. З точки зору розпаду (катаболізму), амінокислоти класифікуються як глюкогенні, якщо вони виробляють проміжні продукти, які можуть бути перетворені в глюкозу або кетогенні, якщо їх проміжні продукти перетворюються в ацетил-КоА. На малюнку 2.13 показані метаболічні долі катаболізму кожної з амінокислот. Відзначимо, що деякі амінокислоти є як глюкогенними, так і кетогенними.
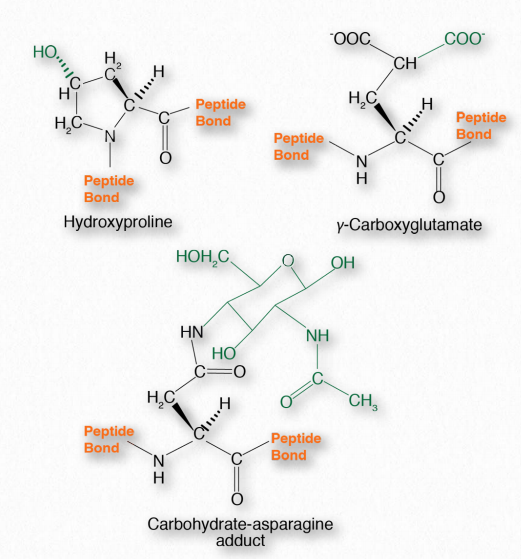
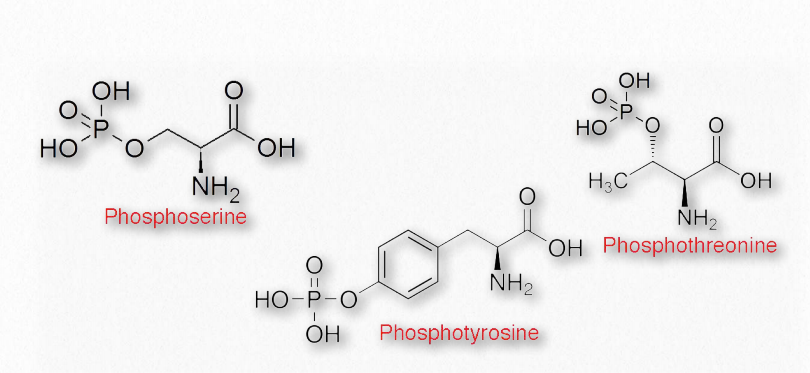
Посттрансляційні модифікації
Після синтезу білка бічні ланцюги амінокислот всередині нього можуть бути хімічно модифіковані, що призводить до більшої різноманітності структури і функції (рис. 2.14). Поширені зміни включають фосфорилювання гідроксильних груп серину, треоніну або тирозину. Лізин, пролін та гістидин можуть мати гідроксили, додані до амінів у їх R-групах. Інші модифікації амінокислот в білках включають додавання жирних кислот (міристинової кислоти або пальмітинової кислоти), ізопреноїдних груп, ацетильних груп, метильних груп, йоду, карбоксильних груп або сульфатів. Вони можуть мати ефекти іонізації (додавання фосфатів/сульфатів), деіонізації (додавання ацетильної групи до R-групи аміну лізину) або взагалі не впливати на заряд. Крім того, N-зв'язані- і О-зв'язаніглікопротеїни мають вуглеводи, ковалентно прикріплені до бічних ланцюгів аспарагіну і треоніну або серину відповідно.
Деякі амінокислоти є попередниками важливих сполук в організмі. Приклади включають адреналін, гормони щитовидної залози, Ldopa та дофамін (все з тирозину), серотонін (з триптофану) та гістамін (з гістидину).
Будівельні поліпептиди
Хоча амінокислоти виконують інші функції в клітині, їх найважливіша роль як складових білків. Білки, як ми відзначали раніше, є полімерами амінокислот.
Амінокислоти пов'язані між собою пептидними зв'язками, в яких карбоксильна група однієї амінокислоти приєднується до аміногрупи наступної, з втратою молекули води. Додаткові амінокислоти додаються таким же чином, шляхом утворення пептидних зв'язків між вільним карбоксилом на кінці зростаючого ланцюга і аміногрупою наступної амінокислоти в послідовності. Ланцюг, що складається лише з декількох амінокислот, пов'язаних між собою, називається олігопептидом (оліго = мало), тоді як типовий білок, який складається з багатьох амінокислот, називається поліпептидом (полі= багато). Кінець пептиду, який має вільну аміногрупу, називається N-кінцевим (для NH2), тоді як кінець з вільним карбоксилом називається C-кінцевим (для карбоксилу).
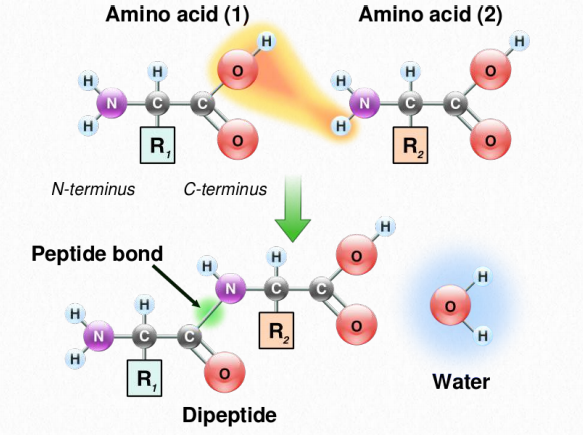
Як ми вже зазначали раніше, функція залежить від структури, і рядок амінокислот повинна складатися в певну 3-D форму, або конформацію, щоб зробити функціональний білок. Згортання поліпептидів в їх функціональні форми - тема наступного розділу.