8.2: катаболізм вуглеводів
- Page ID
- 3945

Цілі навчання
- Опишіть, чому гліколіз не залежить від кисню
- Визначте та опишіть чистий вихід тривуглецевих молекул, АТФ та НАДГ від гліколізу
- Поясніть, як тривуглецеві молекули пірувату перетворюються в двовуглецеві ацетильні групи, які можуть бути спрямовані в цикл Кребса.
- Визначте та опишіть чистий вихід CO 2, GTP/ATP, FADH 2 та NADH з циклу Кребса
- Поясніть, як проміжні молекули вуглецю циклу Кребса можуть бути використані в клітці
Існують великі ферментні шляхи для розщеплення вуглеводів для захоплення енергії в зв'язках АТФ. Крім того, багато катаболічних шляхів виробляють проміжні молекули, які також використовуються як будівельні блоки для анаболізму. Розуміння цих процесів важливо з кількох причин. По-перше, оскільки основні метаболічні процеси є загальними для широкого кола хемогетеротрофних організмів, ми можемо багато чого дізнатися про метаболізм людини, вивчаючи метаболізм у більш легко маніпульованих бактерій, таких як кишкова паличка. По-друге, оскільки збудники тварин та людини також є хемогетеротрофами, вивчення деталей метаболізму у цих бактерій, включаючи можливі відмінності між бактеріальними та людськими шляхами, корисно для діагностики патогенів, а також для виявлення антимікробної терапії, орієнтованої специфічні збудники. Нарешті, вивчення конкретно про шляхи, що беруть участь у хемогетеротрофному метаболізмі, також служить основою для порівняння інших більш незвичайних метаболічних стратегій, що використовуються мікробами. Хоча хімічне джерело електронів, що ініціюють перенесення електронів, відрізняється між хемогетеророфами і хемоавтотрофами, багато подібних процесів використовуються в обох типах організмів.
Типовим прикладом, який використовується для введення учнів понять метаболізму, є вуглеводний катаболізм. Для хемогетеротрофів наші приклади метаболізму починаються з катаболізму полісахаридів, таких як глікоген, крохмаль або целюлоза. Такі ферменти, як амілаза, яка розщеплює глікоген або крохмаль, і целюлази, які розщеплюють целюлозу, можуть викликати гідроліз глікозидних зв'язків між мономерами глюкози в цих полімерах, виділяючи глюкозу для подальшого катаболізму.
гліколіз
Для бактерій, еукаріотів та більшості архей гліколіз є найпоширенішим шляхом катаболізму глюкози; він виробляє енергію, зменшують носії електронів та молекули попередників клітинного метаболізму. Кожен живий організм здійснює якусь форму гліколізу, припускаючи, що цей механізм є древнім універсальним обмінним процесом. Сам процес не використовує кисень, однак гліколіз може поєднуватися з додатковими обмінними процесами, які є або аеробними, або анаеробними. Гліколіз протікає в цитоплазмі прокаріотичних і еукаріотичних клітин. Він починається з однієї шестивуглецевої молекули глюкози і закінчується двома молекулами тривуглецевого цукру під назвою піруват. Піруват може бути розщеплений далі після гліколізу, щоб отримати більше енергії за допомогою аеробного або анаеробного дихання, але багато організмів, включаючи багато мікробів, можуть бути не в змозі дихати; для цих організмів гліколіз може бути їх єдиним джерелом генерації АТФ.
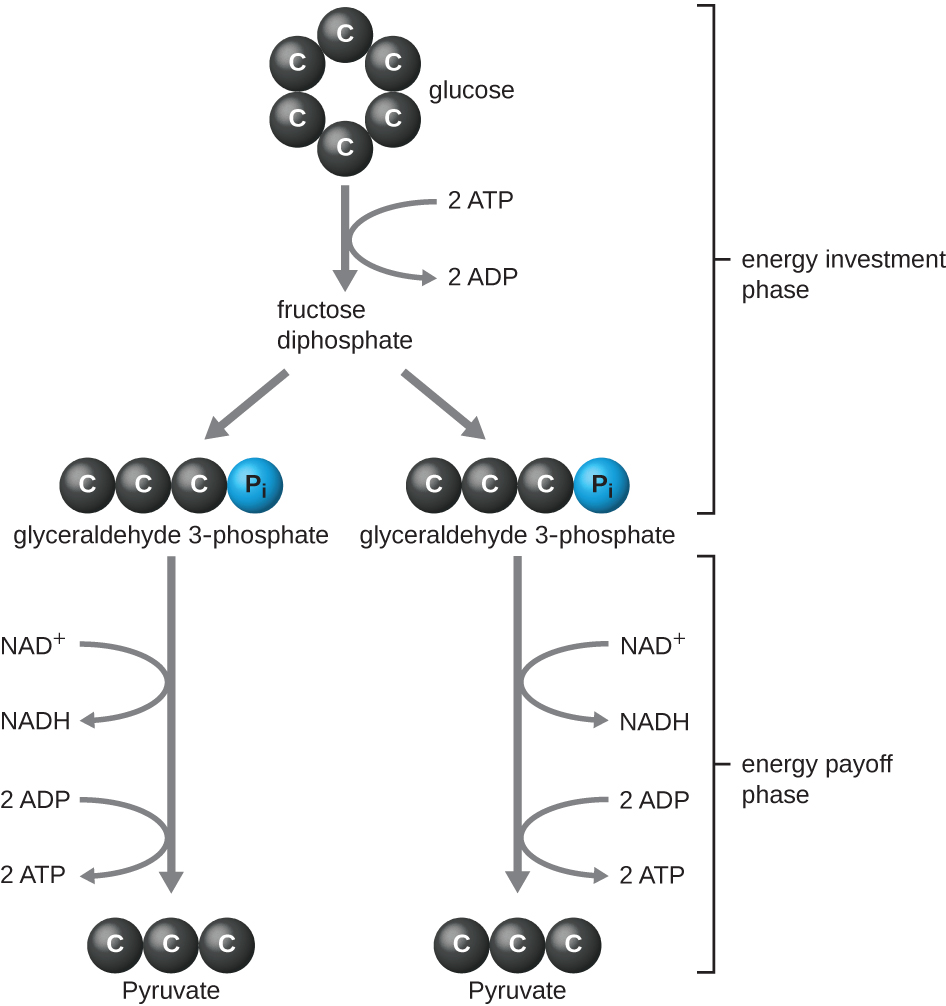
Тип гліколізу, виявленого у тварин і який найбільш поширений у мікробів, - шлях Емден-Мейєргофа-Парнас (EMP), названий на честь Густава Ембдена (1874—1933), Отто Мейєргофа (1884—1951) та Якуба Парнаса (1884—1949). Гліколіз за допомогою шляху EMP складається з двох різних фаз (рис.\(\PageIndex{1}\)). Перша частина шляху, яка називається фазою інвестування енергії, використовує енергію двох молекул АТФ для модифікації молекули глюкози, щоб шестивуглецеву молекулу цукру можна було рівномірно розділити на дві фосфорильовані тривуглецеві молекули, звані гліцеральдегідом 3-фосфат (G3P). Друга частина шляху, яка називається фазою віддачі енергії, витягує енергію шляхом окислення G3P до пірувату, виробляючи чотири молекули АТФ і зменшуючи дві молекули NAD + до двох молекул NADH, використовуючи електрони, що виникли з глюкози. (Обговорення та ілюстрація повного шляху EMP з хімічними структурами та назвами ферментів відображаються в Додатку С.)
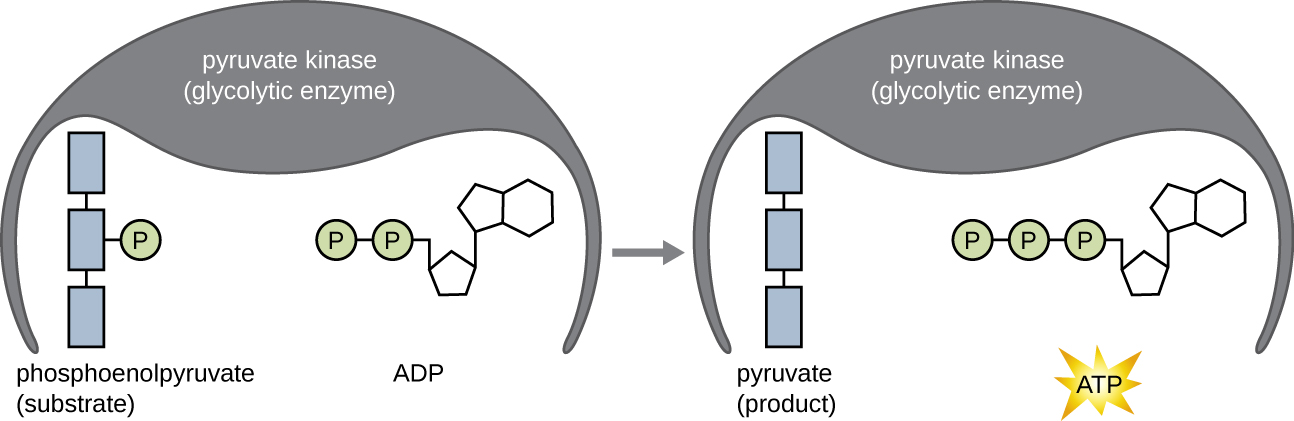
Молекули АТФ, що утворюються під час фази енерговіддачі гліколізу, утворюються фосфорилуванням на рівні субстрату (рис.\(\PageIndex{1}\)), одним з двох механізмів отримання АТФ. При фосфорилюванні на рівні субстрату фосфатна група видаляється з органічної молекули і безпосередньо переноситься в наявну молекулу АДФ, виробляючи АТФ. Під час гліколізу до АДФ додають високоенергетичні фосфатні групи з проміжних молекул, щоб зробити АТФ.
В цілому, в цьому процесі гліколізу чистий приріст від розпаду однієї молекули глюкози становить:
- дві молекули АТФ
- дві молекули NADH, і
- дві молекули пірувату.
Інші гліколітичні шляхи
Коли ми маємо на увазі гліколіз, якщо не вказано інше, ми маємо на увазі шлях EMP, який використовується тваринами та багатьма бактеріями. Однак деякі прокаріоти використовують альтернативні гліколітичні шляхи. Однією з важливих альтернатив є шлях Ентер-Дудорофф (ED), названий на честь його першовідкривачів Натана Ентнера та Майкла Дудорофа (1911—1975). Хоча деякі бактерії, включаючи умовно-патогенний грамнегативний збудник Pseudomonas aeruginosa, містять лише шлях ЕД для гліколізу, інші бактерії, такі як кишкова паличка, мають можливість використовувати або шлях ЕД, або шлях EMP.
Третім типом гліколітичного шляху, який зустрічається у всіх клітині, який сильно відрізняється від попередніх двох шляхів, є пентозно-фосфатний шлях (PPP), який також називають фосфоглюконатним шляхом або шунтом гексози монофосфату. Докази свідчать про те, що ППС може бути найдавнішим універсальним гліколітичним шляхом. Проміжні продукти з ППС використовуються для біосинтезу нуклеотидів і амінокислот. Тому цей гліколітичний шлях може бути сприятливим, коли клітина потребує нуклеїнової кислоти та/або синтезу білка відповідно. Обговорення та ілюстрація повного шляху ЕД та PPP з хімічними структурами та назвами ферментів з'являються у Додатку С.
Вправа\(\PageIndex{1}\)
Коли організм може використовувати шлях ЕД або ППС для гліколізу?
Реакція переходу, коензим А та цикл Кребса
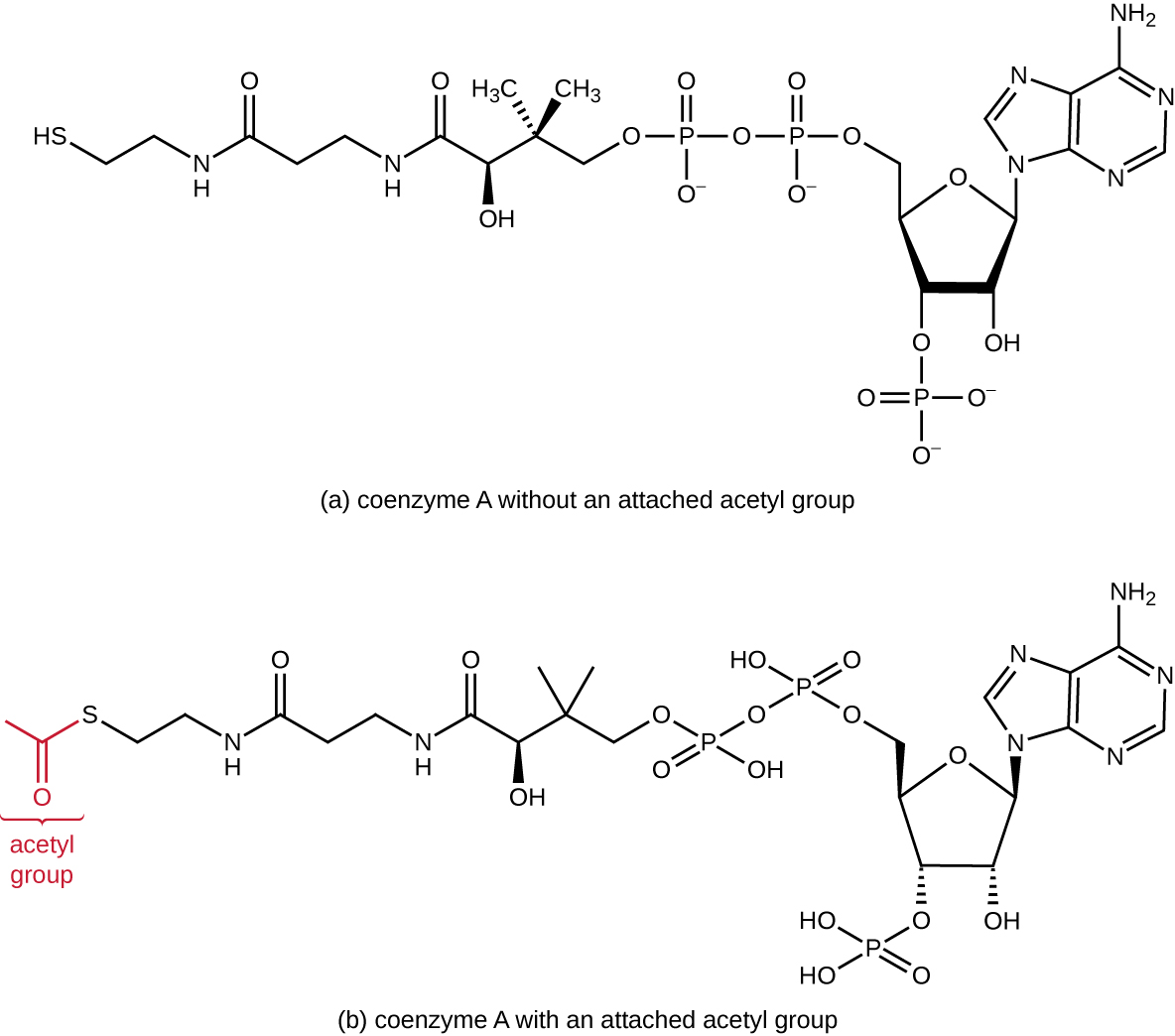
Гліколіз виробляє піруват, який можна додатково окислювати, щоб захопити більше енергії. Щоб піруват потрапив у наступний окислювальний шлях, його спочатку потрібно декарбоксилювати ферментним комплексом піруватдегідрогенази до двовуглецевої ацетильної групи в реакції переходу, яку також називають мостовою реакцією (див. Додаток С та рис.\(\PageIndex{3}\)). У реакції переходу електрони також переносяться в NAD + з утворенням NADH. Щоб перейти до наступної фази цього метаболічного процесу, порівняно крихітний двовуглецевий ацетил повинен бути приєднаний до дуже великої сполуки-носія, яка називається коензимом А (CoA). Реакція переходу відбувається в мітохондріальному матриксі еукаріот; у прокаріотів вона виникає в цитоплазмі, оскільки у прокаріотів відсутні мембрано-закриті органели.
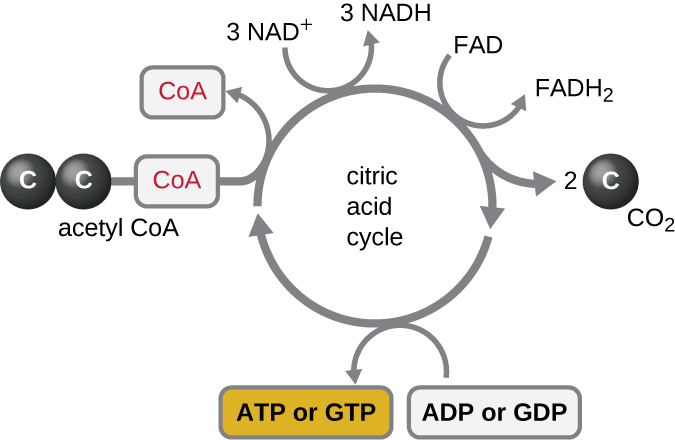
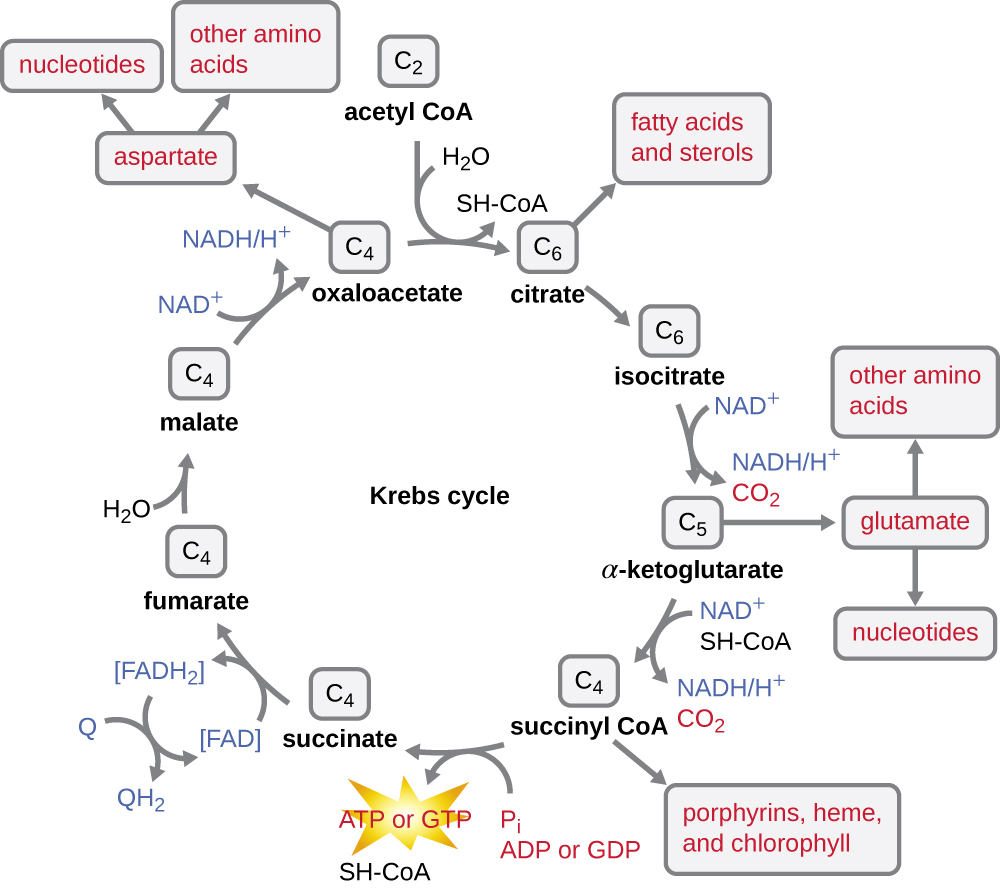
Цикл Кребса переносить залишилися електрони з ацетильної групи, що утворюються в ході реакції переходу, на молекули носія електронів, тим самим зменшуючи їх. Цикл Кребса також відбувається в цитоплазмі прокаріотів разом з гліколізом і реакцією переходу, але він відбувається в мітохондріальному матриксі еукаріотичних клітин, де також відбувається реакція переходу. Цикл Кребса названий на честь його першовідкривача, британського вченого Ганса Адольфа Кребса (1900—1981) і його також називають циклом лимонної кислоти, або циклом трикарбонової кислоти (TCA), оскільки лимонна кислота має три карбоксильні групи у своїй структурі. На відміну від гліколізу, цикл Кребса є замкнутим циклом: Остання частина шляху регенерує з'єднання, яке використовується на першому кроці (рис.\(\PageIndex{4}\)). Вісім етапів циклу являють собою ряд хімічних реакцій, які захоплюють двовуглецеву ацетилгрупу (носій CoA не входить в цикл Кребса) з реакції переходу, яка додається до чотиривуглецевого проміжного продукту в циклі Кребса, виробляючи шестивуглецеву проміжну лимонну кислоту (даючи альтернативне ім'я для цього циклу). Коли один оборот циклу повертається до вихідної точки чотиривуглецевого проміжного продукту, цикл виробляє дві молекули CO 2, одну молекулу АТФ (або еквівалент, наприклад, гуанозинтрифосфат [GTP]), що виробляється фосфорилуванням на рівні субстрату, і три молекули NADH і одну з FADH 2 . (Обговорення та детальна ілюстрація повного циклу Кребса наведено у Додатку C.)
Хоча багато організмів використовують цикл Кребса, описаний як частина метаболізму глюкози, кілька проміжних сполук у циклі Кребса можуть бути використані для синтезу широкого спектру важливих клітинних молекул, включаючи амінокислоти, хлорофіли, жирні кислоти та нуклеотиди; отже, цикл як анаболічні, так і катаболічні (рис.\(\PageIndex{5}\)).
Ключові поняття та резюме
- Гліколіз є першим етапом розпаду глюкози, в результаті чого утворюється АТФ, який виробляється фосфорилювання на рівні субстрату; NADH; і дві молекули пірувату. Гліколіз не використовує кисень і не залежить від кисню.
- Після гліколізу тривуглецевий піруват декарбоксилюють з утворенням двовуглецевої ацетильної групи, поєднаної з утворенням НАДГ. Ацетильна група приєднується до великої сполуки-носія під назвою коензим А.
- Після перехідного етапу коензим А транспортує двовуглецевий ацетил в цикл Кребса, де два вуглеці входять в цикл. За поворот циклу одна ацетильна група, отримана від гліколізу, додатково окислюється, утворюючи три молекули NADH, одну FADH 2 та одну АТФ фосфорилювання на рівні субстрату та вивільняючи дві молекули CO 2.
- Цикл Кребса можна використовувати для інших цілей. Багато проміжних продуктів використовуються для синтезу важливих клітинних молекул, включаючи амінокислоти, хлорофіли, жирні кислоти та нуклеотиди.