8.3: Клітинне дихання
- Page ID
- 3934

Цілі навчання
- Порівняйте та порівняйте розташування та функцію електронної транспортної системи в прокаріотичній клітині та еукаріотичній клітині
- Порівняйте та порівняйте відмінності між рівнем субстрату та окислювальним фосфорилюванням
- Поясніть взаємозв'язок між хеміосмосом та рушійною силою протона
- Опишіть функцію та розташування АТФ-синтази в прокаріотичній та еукаріотичній клітині
- Порівняйте і контрастуйте аеробне і анаеробне дихання
Ми щойно обговорили два шляхи катаболізму глюкози - гліколіз та цикл Кребса - які генерують АТФ шляхом фосфорилювання на рівні субстрату. Більшість АТФ, однак, генерується під час окремого процесу, який називається окислювальним фосфорилуванням, який виникає під час клітинного дихання. Клітинне дихання починається, коли електрони передаються від NADH та FADH 2 - зроблені в гліколізі, реакції переходу та циклі Кребса - через ряд хімічних реакцій до кінцевого неорганічного акцептора електронів (або кисень в аеробному диханні, або некисневі неорганічні молекули в анаеробне дихання). Ці перенесення електронів відбуваються на внутрішній частині клітинної мембрани прокаріотичних клітин або в спеціалізованих білкових комплексах у внутрішній мембрані мітохондрій еукаріотичних клітин. Енергія електронів збирається для генерації електрохімічного градієнта через мембрану, який використовується для отримання АТФ шляхом окислювального фосфорилювання.
Електронна транспортна система
Електронно-транспортна система (ЕТС) є останнім компонентом, який бере участь у процесі клітинного дихання; вона містить ряд мембранасоційованих білкових комплексів та асоційованих мобільних допоміжних носіїв електронів. Електронний транспорт - це серія хімічних реакцій, що нагадує бригаду ковша в тому, що електрони від NADH і FADH 2 швидко передаються від одного електронного носія ETS до іншого. Ці носії можуть пропускати електрони вздовж в ETS через їх окислювально-відновного потенціалу. Щоб білок або хімікат приймав електрони, він повинен мати більш позитивний окислювально-відновний потенціал, ніж донор електронів. Тому електрони рухаються від електронних носіїв з більшим негативним окислювально-відновним потенціалом до тих, у кого більше позитивного окислювально-відновного потенціалу. Чотири основні класи електронних носіїв, що беруть участь як в еукаріотичних, так і в прокаріотичних системах електронного транспорту, - це цитохроми, флавопротеїни, залізо-сірчані білки та хінони.
При аеробному диханні кінцевий акцептор електронів (тобто той, що має найбільш позитивний окислювально-відновний потенціал) в кінці ETS - це молекула кисню (O 2), яка зменшується до води (H 2 O) кінцевим носієм ETS. Цей носій електронів, цитохромоксидаза, відрізняється між типами бактерій і може бути використаний для диференціації тісно пов'язаних бактерій для діагнозів. Наприклад, грамнегативний опортуніст Pseudomonas aeruginosa та грамнегативні холери Vibrio cholerae використовують цитохром c оксидазу, яку можна виявити за допомогою тесту на оксидазу, тоді як інші грамнегативні Enterobacteriaceae, як E. coli, є негативними для цього тест, оскільки вони виробляють різні типи цитохромоксидази.
Існує багато обставин, при яких аеробне дихання неможливе, включаючи будь-яке одне або кілька з наступних:
- Клітині не вистачає генів, що кодують відповідну цитохромоксидазу для перенесення електронів кисню в кінці електронної транспортної системи.
- Клітині не вистачає генів, що кодують ферменти, щоб мінімізувати сильно руйнівний вплив небезпечних кисневих радикалів, що утворюються під час аеробного дихання, таких як перекис водню (H 2 O 2) або супероксид\((\ce{O2-})\).
- Клітці не вистачає достатньої кількості кисню для здійснення аеробного дихання.
Однією з можливих альтернатив аеробному диханню є анаеробне дихання, використовуючи неорганічну молекулу, відмінну від кисню, як кінцевий акцептор електронів. Існує багато типів анаеробного дихання, виявлених у бактерій та архей. Денітрифікатори є важливими ґрунтовими бактеріями, які використовують нітрати\((\ce{NO3-})\) та нітрити\((\ce{NO2-})\) як кінцеві акцептори електронів, виробляючи газ азоту (N 2). Багато аеробно дихальних бактерій, включаючи кишкову паличку, переходять на використання нітрату як кінцевого акцептора електронів і виробляють нітрит, коли рівень кисню був виснажений.
Мікроби, що використовують анаеробне дихання, зазвичай мають інтактний цикл Кребса, тому ці організми можуть отримати доступ до енергії утворених молекул NADH і FADH 2. Однак анаеробні респіратори використовують змінені носії ETS, закодовані їх геномами, включаючи різні комплекси для перенесення електронів до їх кінцевих акцепторів електронів. Менші електрохімічні градієнти генеруються з цих систем перенесення електронів, тому менше АТФ утворюється через анаеробне дихання.
Вправа\(\PageIndex{1}\)
Чи використовують як аеробне дихання, так і анаеробне дихання ланцюг транспорту електронів?
Хіміосмос, рушійна сила протонів та окислювальне фосфорилювання
При кожному перенесенні електрона через ETS електрон втрачає енергію, але при деяких передачах енергія зберігається як потенційна енергія, використовуючи її для перекачування іонів водню (H +) через мембрану. У прокаріотичних клітині Н + перекачується назовні цитоплазматичної мембрани (у грамнегативних і грампозитивних бактерій називається периплазматичним простором), а в еукаріотичних клітині вони перекачуються з мітохондріального матриксу через внутрішню мембрану мітохондрій в міжмембранний простір. Існує нерівномірний розподіл Н + по мембрані, що встановлює електрохімічний градієнт, оскільки іони H + позитивно заряджені (електричні) і є більш висока концентрація (хімічна) на одній стороні мембрани. Цей електрохімічний градієнт, утворений накопиченням H + (також відомого як протон) на одній стороні мембрани порівняно з іншою, називається рушійною силою протона (PMF). Оскільки беруть участь іони H +, також встановлюється градієнт рН, причому сторона мембрани, що має більш високу концентрацію Н +, є більш кислою. Окрім використання ПМФ для виготовлення АТФ, як обговорюється в цьому розділі, ПМФ також може використовуватися для керування іншими енергетично несприятливими процесами, включаючи транспорт поживних речовин та обертання джгутиків для рухливості.
Потенційна енергія цього електрохімічного градієнта, що генерується ETS, змушує H + дифузно через мембрану (плазматичну мембрану в прокаріотичних клітині та внутрішню мембрану в мітохондріях в еукаріотичних клітині). Цей потік іонів водню через мембрану, званий хеміосмосом, повинен відбуватися через канал в мембрані через мембранно-зв'язаний ферментний комплекс під назвою АТФ-синтаза (рис.\(\PageIndex{1}\)). Тенденція руху таким чином багато в чому схожа на воду, що скупчилася з одного боку греблі, що рухається через дамбу при відкритті. АТФ-синтаза (як комбінація забору і генератора гідроелектричної греблі) являє собою складний білок, який діє як крихітний генератор, повертаючись силою H +, дифузірующій через фермент, вниз їх електрохімічний градієнт звідки багато взаємно відштовхують Н + до де менше Н +. У прокаріотичних клітинок Н + перетікає з зовнішньої сторони цитоплазматичної мембрани в цитоплазму, тоді як в еукаріотичних мітохондріях Н + протікає з міжмембранного простору в мітохондріальний матрикс. Поворот частин цієї молекулярної машини регенерує АТФ з АДФ та неорганічного фосфату (P i) шляхом окислювального фосфорилювання, другого механізму виготовлення АТФ, який збирає потенційну енергію, що зберігається в електрохімічному градієнті.
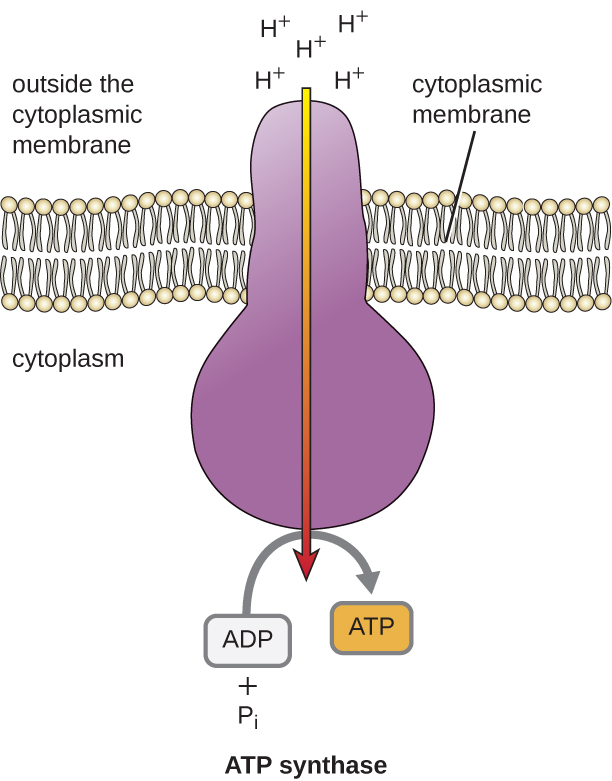
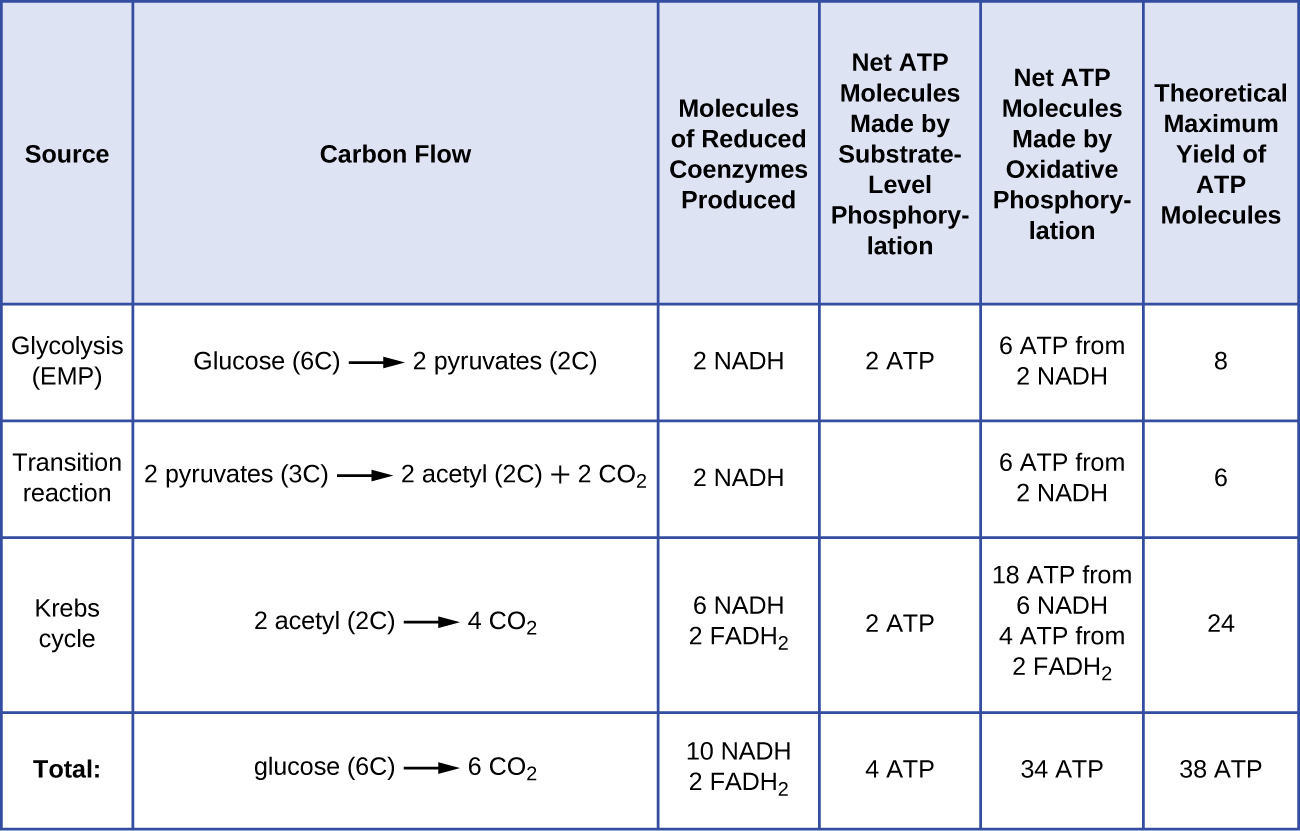
Кількість молекул АТФ, що утворюються в результаті катаболізму глюкози, змінюється. Наприклад, кількість іонів водню, які комплекси електронної транспортної системи можуть прокачувати через мембрану, коливається між різними видами організмів. При аеробному диханні в мітохондріях проходження електронів з однієї молекули NADH генерує достатню протонну рушійну силу, щоб зробити три молекули АТФ шляхом окислювального фосфорилювання, тоді як проходження електронів з однієї молекули FADH 2 генерує достатню протонну рушійну силу, щоб зробити лише дві Молекули АТФ. Таким чином, 10 молекул NADH, виготовлених на глюкозу під час гліколізу, реакції переходу та циклу Кребса, несуть достатньо енергії, щоб зробити 30 молекул АТФ, тоді як дві молекули FADH 2, виготовлені на глюкозу під час цих процесів, забезпечують достатньо енергії, щоб зробити чотири молекули АТФ. Загалом, теоретичний максимальний вихід АТФ, зроблений під час повного аеробного дихання глюкози, становить 38 молекул, причому чотири - фосфорилювання на рівні субстрату, а 34 - шляхом окислювального фосфорилювання (рис.\(\PageIndex{2}\)). Насправді загальний вихід АТФ зазвичай менше, починаючи від однієї до 34 молекул АТФ, залежно від того, чи використовує клітина аеробне дихання або анаеробне дихання; в еукаріотичних клітині частина енергії витрачається на транспортування проміжних продуктів з цитоплазми в мітохондрії, впливаючи на вихід АТФ.
Малюнок\(\PageIndex{2}\) узагальнює теоретичні максимальні виходи АТФ від різних процесів при повному аеробному диханні однієї молекули глюкози.
Вправа\(\PageIndex{1}\)
Які функції має рушійна сила протона?
Резюме
- Більшість АТФ, що утворюється під час клітинного дихання глюкози, виробляється шляхом окислювального фосфорилювання.
- Електронно-транспортна система (ETS) складається з ряду мембранно-асоційованих білкових комплексів та пов'язаних з ними мобільних допоміжних електронних носіїв. ЕТС вбудовується в цитоплазматичну мембрану прокаріотів і внутрішню мітохондріальну мембрану еукаріотів.
- Кожен комплекс ETS має різний окислювально-відновний потенціал, і електрони рухаються від електронних носіїв з більшим негативним окислювально-відновним потенціалом до тих, у кого більше позитивного окислювально-відновного потенціалу.
- Для здійснення аеробного дихання клітці потрібен кисень як кінцевий акцептор електронів. Клітині також потрібен повний цикл Кребса, відповідна цитохромоксидаза та ферменти детоксикації кисню, щоб запобігти шкідливому впливу кисневих радикалів, що утворюються під час аеробного дихання.
- Організми, які виконують анаеробне дихання, використовують альтернативні носії електронної транспортної системи для остаточного перенесення електронів до кінцевих некисневих акцепторів електронів.
- Мікроби виявляють великі варіації в складі своїх електронних транспортних систем, які можуть бути використані в діагностичних цілях, щоб допомогти виявити певні збудники.
- Коли електрони передаються від NADH і FADH 2 через ETS, електрон втрачає енергію. Ця енергія накопичується за допомогою накачування Н + через мембрану, генеруючи протонну рушійну силу.
- Енергія цієї рушійної сили протона може бути використана, дозволяючи іонам водню дифузувати назад через мембрану шляхом хеміосмосу з використанням АТФ-синтази. Оскільки іони водню дифундують через їх електрохімічний градієнт, компоненти АТФ-синтази обертаються, роблячи АТФ з АДФ і P i шляхом окислювального фосфорилювання.
- Аеробне дихання утворює більше АТФ (максимум 34 молекули АТФ) під час окислювального фосфорилювання, ніж анаеробне дихання (між однією і 32 молекулами АТФ).