3.3: Унікальні характеристики прокаріотичних клітин
- Page ID
- 4121

Цілі навчання
- Поясніть відмінні характеристики прокаріотичних клітин
- Опишіть загальні морфології клітин і клітинні композиції, характерні для прокаріотичних клітин, і поясніть, як клітини підтримують свою морфологію
- Охарактеризуйте внутрішні та зовнішні структури прокаріотичних клітин з точки зору їх фізичної структури, хімічної структури та функції
- Порівняйте відмінні характеристики бактеріальних і архейних клітин
Клітинна теорія стверджує, що клітина є фундаментальною одиницею життя. Однак клітини значно різняться за розміром, формою, структурою та функцією. На найпростішому рівні побудови всі клітини володіють декількома основними складовими. До них відносяться цитоплазма (гелеподібна речовина, що складається з води та розчинених хімічних речовин, необхідних для росту), яка міститься в плазматичній мембрані (також називається клітинною мембраною або цитоплазматичною мембраною); одна або кілька хромосом, які містять генетичні креслення клітини; і рибосоми, органели, використовувані для виробництва білків.
Крім цих основних компонентів, клітини можуть сильно відрізнятися між організмами і навіть в межах одного і того ж багатоклітинного організму. Дві найбільші категорії клітин - прокаріотичні клітини та еукаріотичні клітини - визначаються основними відмінностями в декількох клітинних структурах. Прокаріотичні клітини не мають ядра, оточеного складною ядерною мембраною і, як правило, мають єдину кругову хромосому, розташовану в нуклеоїді. Еукаріотичні клітини мають ядро, оточене складною ядерною мембраною, яка містить множинні, стрижневі хромосоми. 1
Всі рослинні клітини і клітини тварин є еукаріотичними. Деякі мікроорганізми складаються з прокаріотичних клітин, тоді як інші складаються з еукаріотичних клітин. Прокаріотичні мікроорганізми класифікуються в межах областей Архея і Бактерії, тоді як еукаріотичні організми класифікуються в межах області Eukarya.
Структури всередині клітини аналогічні органам всередині людського тіла, з унікальними структурами, придатними для конкретних функцій. Деякі структури, знайдені в прокаріотичних клітині, схожі на ті, що містяться в деяких еукаріотичних клітині; інші унікальні для прокаріотів. Хоча є деякі винятки, еукаріотичні клітини, як правило, більші за прокаріотичні клітини. Порівняно більший розмір еукаріотичних клітин диктує необхідність компартменталізації різних хімічних процесів в межах різних областей клітини, використовуючи складні мембранно-зв'язані органели. На відміну від цього, прокаріотичним клітинам, як правило, не вистачає мембрано-зв'язаних органел; однак вони часто містять включення, які компартментують їх цитоплазму. Малюнок\(\PageIndex{1}\) ілюструє структури, зазвичай пов'язані з прокаріотичними клітинами. Більш докладно ці конструкції описані в наступному розділі.
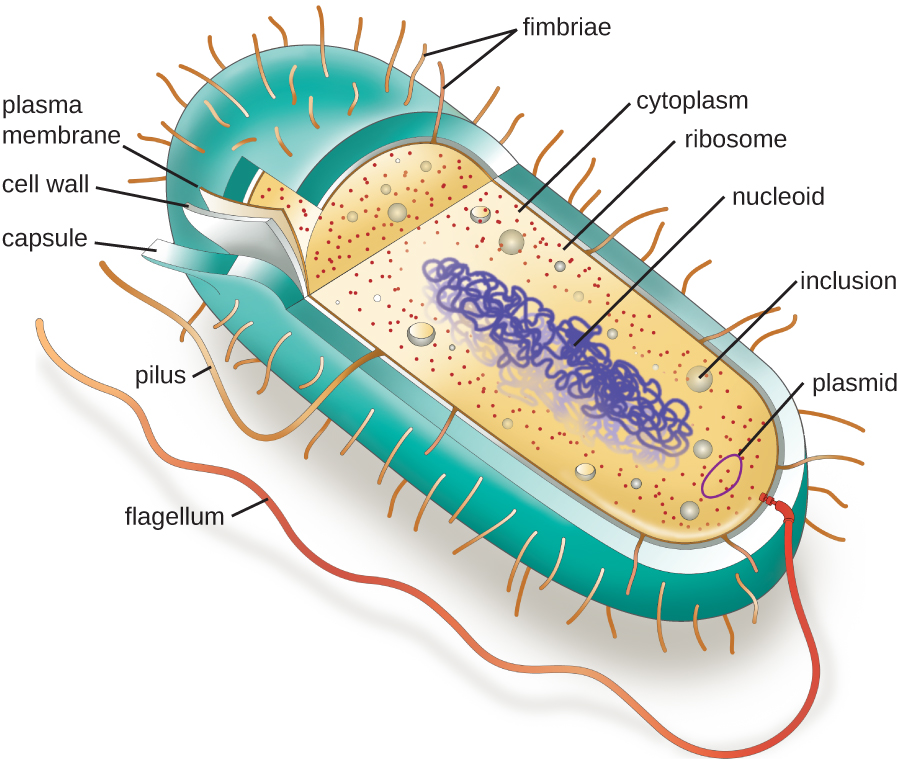
Загальні морфології та розташування клітин
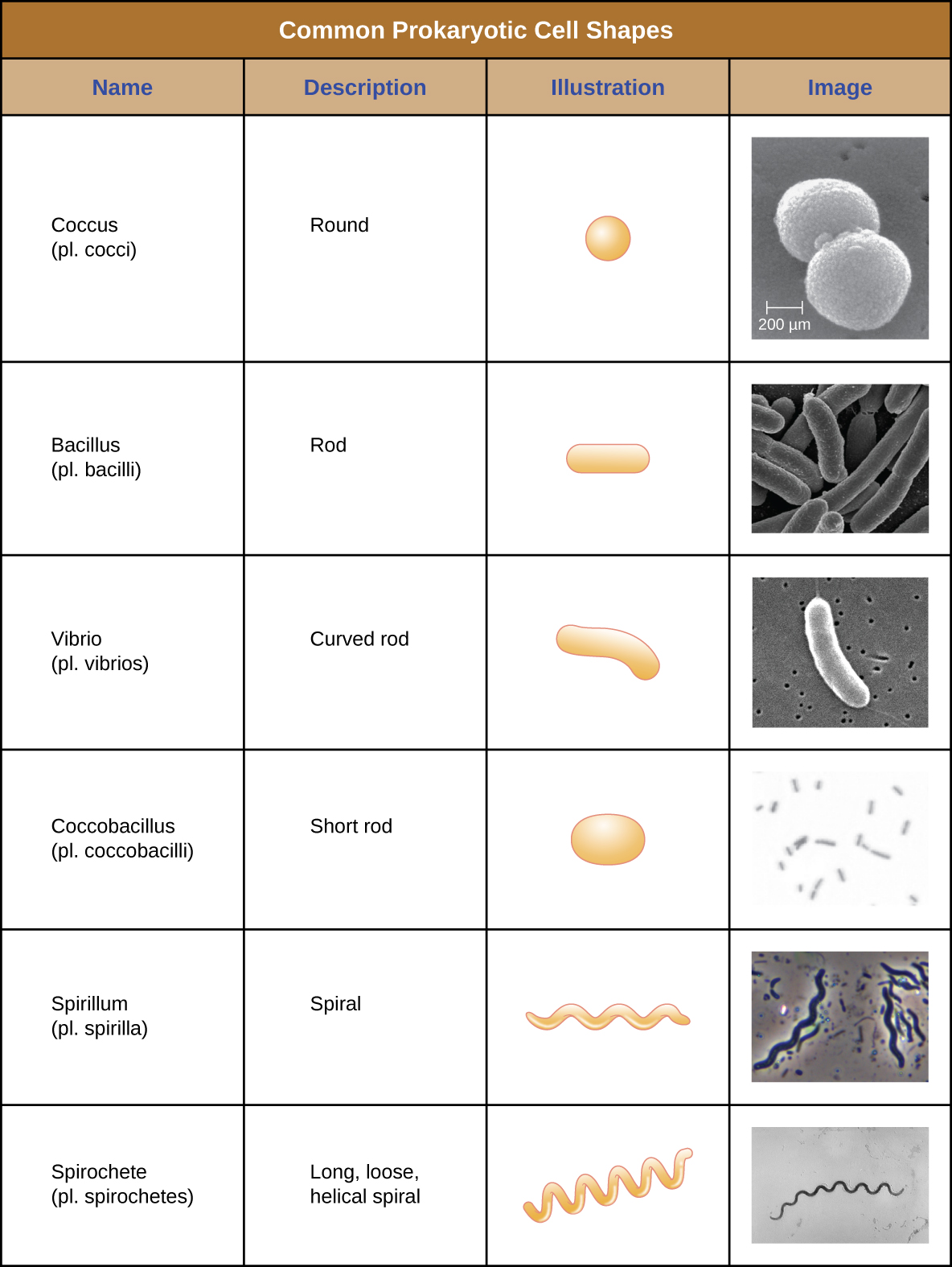
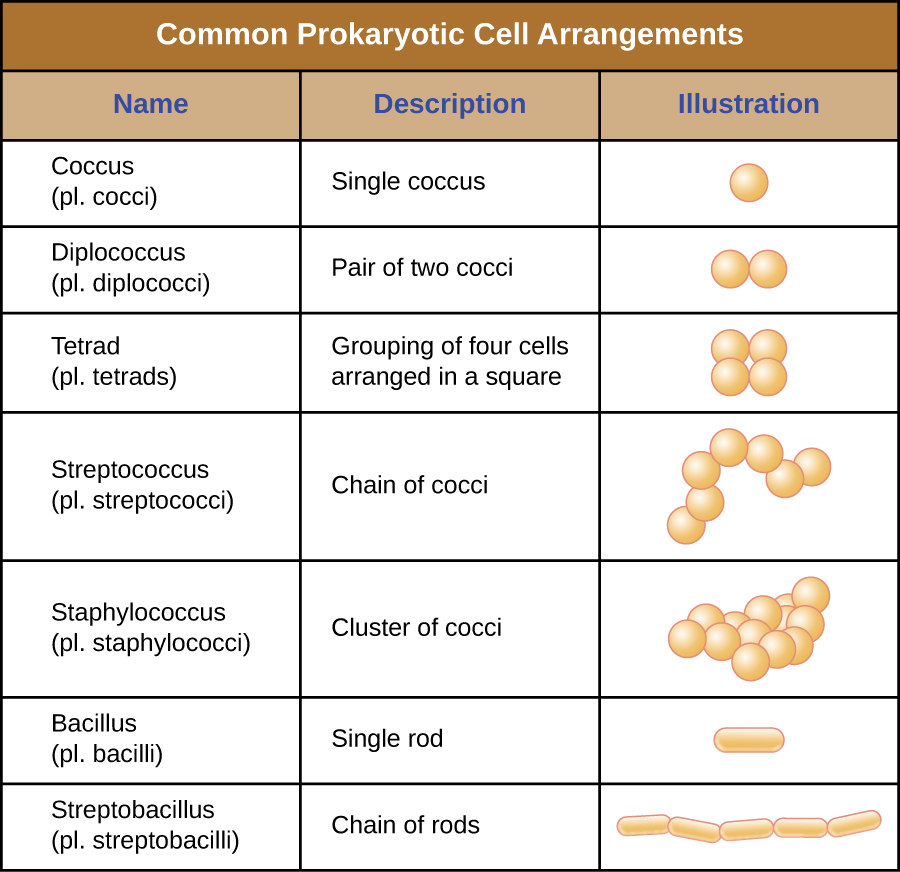
Окремі клітини конкретного прокаріотичного організму, як правило, схожі за формою, або морфологією клітин. Хоча тисячі прокаріотичних організмів були ідентифіковані, лише жменька морфології клітин зазвичай спостерігається мікроскопічно. Малюнок\(\PageIndex{2}\) називає і ілюструє морфологію клітин, які зазвичай зустрічаються в прокаріотичних клітині. На додаток до клітинної форми, прокаріотичні клітини одного виду можуть групуватися в певних відмітних розташуваннях залежно від площини поділу клітин. Деякі загальні домовленості показані на малюнку\(\PageIndex{3}\).
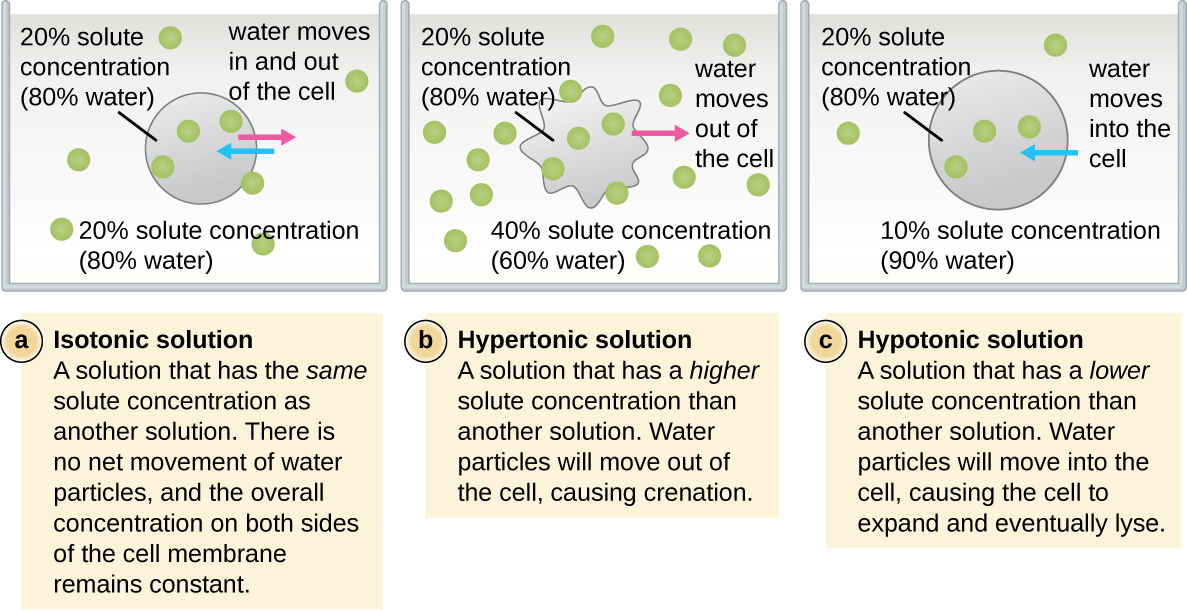
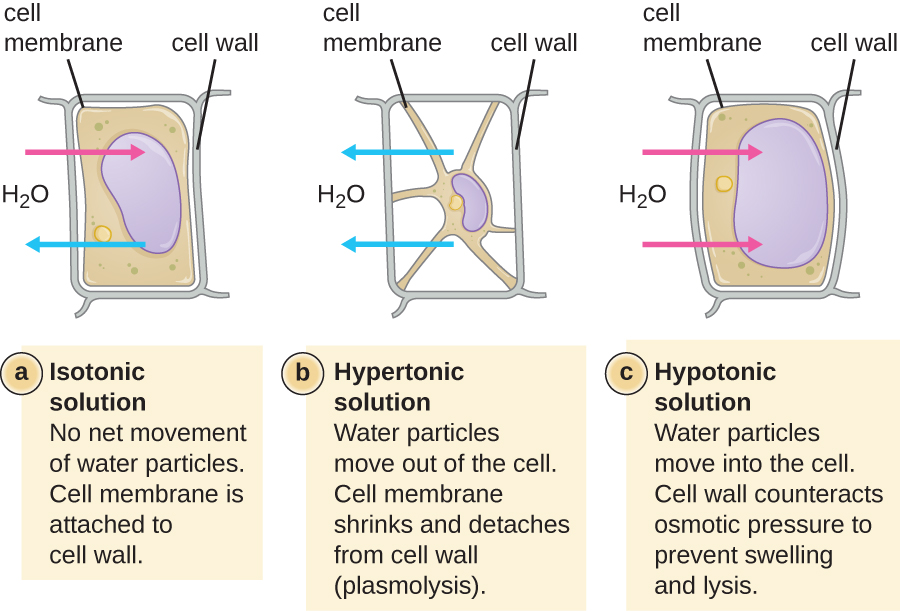
У більшості прокаріотичних клітин морфологія підтримується клітинною стінкою в поєднанні з цитоскелетними елементами. Клітинна стінка - це структура, яка зустрічається у більшості прокаріотів і деяких еукаріотів; вона обволікає клітинну мембрану, захищаючи клітину від змін осмотичного тиску (рис.\(\PageIndex{4}\)). Осмотичний тиск виникає через різницю в концентрації розчинених речовин на протилежних сторонам напівпроникної мембрани. Вода здатна проходити через напівпроникну мембрану, але розчинені речовини (розчинені молекули, такі як солі, цукру та інші сполуки) не можуть. Коли концентрація розчинених речовин більша на одній стороні мембрани, вода дифундує по мембрані з боку з нижчою концентрацією (більше води) в сторону з більш високою концентрацією (менше води), поки концентрації з обох сторін не стануть рівними. Ця дифузія води називається осмосом, і вона може викликати екстремальний осмотичний тиск на клітину при зміні її зовнішнього середовища.
Зовнішнє середовище клітини можна описати як ізотонічну, гіпертонічну або гіпотонічну середовище. В ізотонічному середовищі концентрації розчинених речовин всередині і зовні клітини приблизно рівні, тому немає чистого руху води по клітинній мембрані. У гіпертонічному середовищі концентрація розчиненої речовини поза клітиною перевищує концентрацію всередині клітини, тому вода дифундує з клітини і в зовнішнє середовище. У гіпотонічному середовищі концентрація розчиненої речовини всередині клітини перевищує концентрацію зовні клітини, тому вода буде рухатися осмосом в клітину. Це призводить до того, що клітина набрякає і потенційно лізе, або лопне.
Ступінь, до якої конкретна клітина здатна протистояти перепадам осмотичного тиску, називається тонусом. Клітини, які мають клітинну стінку, краще здатні протистояти тонким змінам осмотичного тиску і зберігати свою форму. У гіпертонічних середовищах клітини, яким не вистачає клітинної стінки, можуть зневоднюватися, викликаючи церенацію або зморщення клітини; плазматична мембрана стискається і виглядає фестончастою або зубчастою (рис.\(\PageIndex{4}\)). На відміну від цього, клітини, які мають клітинну стінку, піддаються плазмолізу, а не крінації. При плазмолізі плазмова мембрана стискається і відривається від клітинної стінки, і відбувається зменшення внутрішнього обсягу, але клітинна стінка залишається неушкодженою, що дозволяє клітині протягом певного періоду часу зберігати певну форму і цілісність (рис.\(\PageIndex{5}\)). Так само клітини, яким не вистачає клітинної стінки, більш схильні до лізису в гіпотонічних середовищах. Наявність клітинної стінки дозволяє клітці довше зберігати свою форму і цілісність перед лізуванням (рис.\(\PageIndex{5}\)).
Вправа\(\PageIndex{1}\)
- Поясніть різницю між морфологією клітин і розташуванням.
- Які переваги надають клітинам прокаріотичних клітин клітинні стінки?
Нуклеоїд
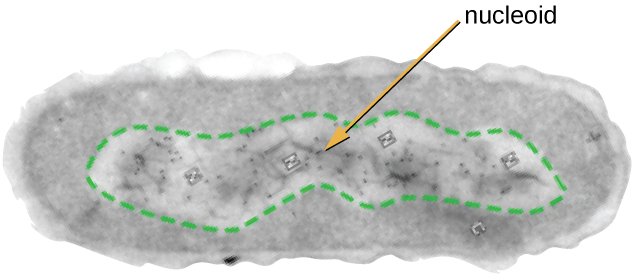
Вся клітинна життя має геном ДНК, організований в одну або кілька хромосом. Прокаріотичні хромосоми, як правило, кругові, гаплоїдні (непарні) і не пов'язані складною ядерною мембраною. Прокаріотична ДНК і ДНК-асоційовані білки зосереджені всередині нуклеоїдної області клітини (рис.\(\PageIndex{6}\)). Взагалі прокаріотична ДНК взаємодіє з нуклеоїдно-асоційованими білками (НПД), які допомагають в організації та упаковці хромосоми. У бактерій НПД функціонують подібно до гістонів, які є дНК-організовуючими білками, що містяться в еукаріотичних клітині. В архей нуклеоїд організовується або НПД, або гістоноподібними ДНК організовуючими білками.
Плазміди
Прокаріотичні клітини також можуть містити екстрахромосомну ДНК, або ДНК, яка не входить до складу хромосоми. Ця екстрахромосомна ДНК міститься в плазмідах, які є невеликими, круговими, дволанцюговими молекулами ДНК. Клітини, які мають плазміди, часто мають сотні їх в межах однієї клітини. Плазміди частіше зустрічаються в бактеріях; однак плазміди були виявлені в архей та еукаріотичних організмах. Плазміди часто несуть гени, які надають вигідні риси, такі як стійкість до антибіотиків; таким чином, вони важливі для виживання організму. Більш детально ми обговоримо плазміди в Механізмах мікробної генетики.
Рибосоми
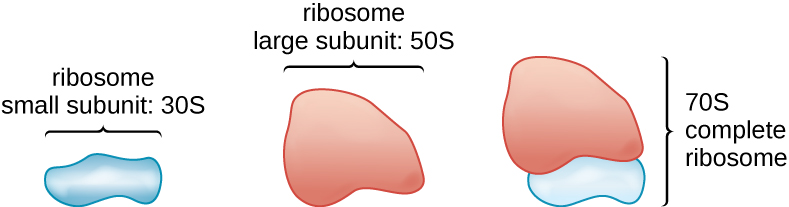
Вся клітинна життя синтезує білки, а організми у всіх трьох областях життя володіють рибосомами, структурами, відповідальними за синтез білка. Однак рибосоми в кожній з трьох областей структурно різні. Самі рибосоми побудовані з білків разом з рибосомальною РНК (рРНК). Прокаріотичні рибосоми виявляються в цитоплазмі. Їх називають рибосомами 70S, оскільки вони мають розмір 70S (рис.\(\PageIndex{7}\)), тоді як еукаріотичні цитоплазматичні рибосоми мають розмір 80S. (S означає одиницю Сведберга, міру осідання в ультрацентрифузі, яка базується на розмірі, формі та поверхневих якостях аналізованої структури). Хоча вони однакового розміру, бактеріальні та архейні рибосоми мають різні білки та молекули рРНК, а архейні версії більше схожі на їх еукаріотичні аналоги, ніж ті, що містяться у бактерій.
Включення
Як одноклітинні організми, що живуть у нестабільних середовищах, деякі прокаріотичні клітини мають здатність зберігати надлишок поживних речовин у цитоплазматичних структурах, які називаються включеннями. Зберігання поживних речовин у полімеризованій формі є вигідним, оскільки воно зменшує накопичення осмотичного тиску, що виникає, коли клітина накопичує розчинені речовини. Різні типи включень зберігають глікоген і крохмалі, які містять вуглець, до якого клітини можуть отримати доступ для отримання енергії. Гранули волютину, які також називають метахроматичними гранулами через їх характеристики фарбування, є включеннями, які зберігають полімеризований неорганічний фосфат, який можна використовувати в обміні речовин і сприяти утворенню біоплівок. Мікроби, як відомо, містять гранули волютину, включають архею метаносарцину, бактерію Corynebacterium diphtheriae та одноклітинну еукаріотичну водорость Chlamydomonas. Гранули сірки, інший тип включення, містяться в сірчаних бактеріях роду Thiobacillus; ці гранули зберігають елементарну сірку, яку бактерії використовують для обміну речовин.
Іноді певні види включень оточені фосфоліпідним моношаром, вбудованим білком. Полігідроксибутират (PHB), який може вироблятися видами Bacillus і Pseudomonas, є прикладом включення, яке відображає цей тип моношарової структури. Промислово PHB також використовується як джерело біологічно розкладаються полімерів для біопластики. Кілька різних видів включень показані на малюнку\(\PageIndex{8}\).

Деякі прокаріотичні клітини мають інші типи включень, які служать іншим цілям, крім зберігання поживних речовин. Наприклад, деякі прокаріотичні клітини виробляють газові вакуолі, скупчення дрібних, вистелених білками бульбашок газу. Ці газові вакуолі дозволяють прокаріотичним клітинам, які їх синтезують, змінювати свою плавучість, щоб вони могли регулювати своє розташування в товщі води. Магнітотаксичні бактерії, такі як Magnetospirillum magnetotacticum, містять магнітосоми, які є включеннями магнітного оксиду заліза або сульфіду заліза, оточених ліпідним шаром. Вони дозволяють клітинам вирівнюватися по магнітному полю, сприяючи їх руху (рис.\(\PageIndex{8}\)). Ціанобактерії, такі як Anabaena cylindrica та бактерії, такі як Halothiobacillus neapolitanus, виробляють карбоксисомні включення. Карбоксисоми складаються з зовнішніх оболонок тисяч білкових субодиниць. Їх внутрішня частина наповнена рибулозою-1,5-бісфосфаткарбоксилазою/оксигеназою (RubiSCo) і карбоангідразою. Обидва ці сполуки використовуються для вуглецевого обміну. Деякі прокаріотичні клітини також мають карбоксисоми, які секвестують функціонально пов'язані ферменти в одному місці. Ці структури вважаються протоорганелами, оскільки вони розділяють важливі сполуки або хімічні реакції, подібно до багатьох еукаріотичних органел.
Ендоспори
Бактеріальні клітини, як правило, спостерігаються як вегетативні клітини, але деякі пологи бактерій мають здатність утворювати ендоспори, структури, які по суті захищають геном бактерій в сплячому стані, коли умови навколишнього середовища несприятливі. Ендоспори (не плутати з репродуктивними спорами, утвореними грибками) дозволяють деяким бактеріальним клітинам довго виживати без їжі або води, а також впливу хімічних речовин, екстремальних температур і навіть радіації. Таблиця\(\PageIndex{1}\) порівнює характеристики вегетативних клітин і ендоспор.
| Вегетативні клітини | Ендоспори |
|---|---|
| Чутливий до екстремальних температур і випромінювання | Стійкий до екстремальних температур і випромінювання |
| грампозитивний | Не вбирають пляму Грама, тільки спеціальні ендоспорові плями (див. Фарбування мікроскопічних зразків) |
| Нормальний вміст води та ферментативна активність | Зневоднений; відсутність метаболічної активності |
| Здатний до активного росту і обміну речовин | Сплячий; відсутність росту або метаболічної активності |
Процес, за допомогою якого вегетативні клітини перетворюються в ендоспори, називається споролюванням, і він, як правило, починається, коли поживні речовини виснажуються або умови навколишнього середовища стають інакше несприятливими (рис.\(\PageIndex{9}\)). Процес починається з утворення перегородки в вегетативної бактеріальної клітці. Перегородка ділить клітину асиметрично, відокремлюючи передміхурову спору ДНК від материнської клітини. Передній спор, який буде утворювати ядро ендоспори, по суті є копією хромосом клітини, і відокремлюється від материнської клітини другою мембраною. Кора поступово утворюється навколо передньої спори, закладаючи між мембранами шари кальцію та дипіколінової кислоти. Потім навколо кори утворюється білкова спорова оболонка, тоді як ДНК материнської клітини розпадається. Подальше дозрівання ендоспори відбувається з утворенням крайньої екзоспорії. Ендоспора звільняється при розпаді материнської клітини, завершуючи спорокуляцію.
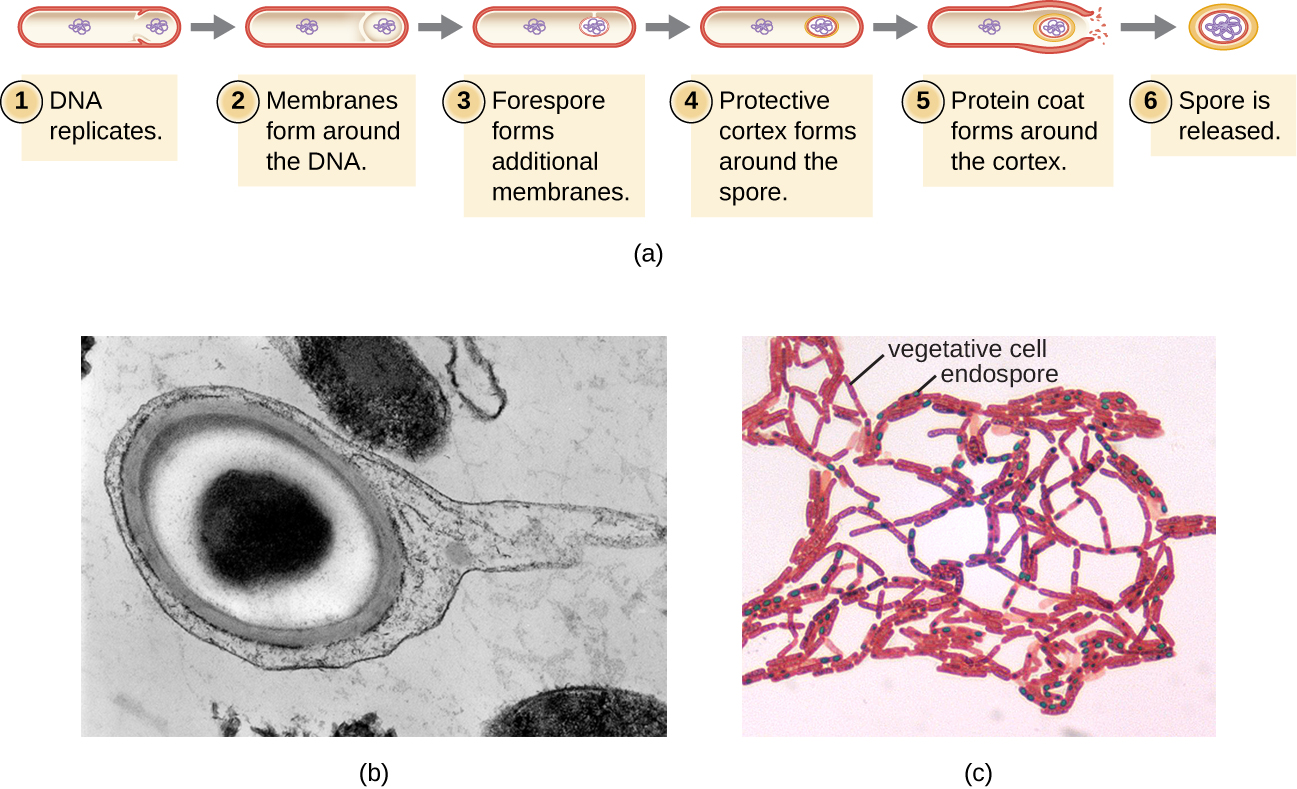
Показано, що ендоспори певних видів зберігаються в стані спокою протягом тривалих періодів часу, до тисяч років. 2 Однак при поліпшенні умов життя ендоспори піддаються проростанню, знову вступаючи в вегетативний стан. Після проростання клітина знову стає метаболічно активною і здатна виконувати всі свої нормальні функції, включаючи ріст і поділ клітин.
Не всі бактерії мають здатність утворювати ендоспори, однак існує ряд клінічно значущих ендоспороутворюючих грампозитивних бактерій пологів Bacillus і Clostridium. До них відносяться B. anthracis, збудник сибірської виразки, який виробляє ендоспори, здатні виживати протягом багатьох десятиліть 3; C. tetani (викликає правець); C. difficile (викликає псевдомембранозний коліт); C. perfringens (викликає газову гангрену); і C. botulinum (викликає ботулізм). Такі патогени, як ці, особливо важко боротися, оскільки їх ендоспори так важко вбити. Спеціальні методи стерилізації ендоспороутворюючих бактерій обговорюються в Контролі росту мікробів.
Вправа\(\PageIndex{2}\)
- Що таке інклюзія?
- Яка функція ендоспори?
Плазмові мембрани
Структури, що охоплюють цитоплазму і внутрішні структури клітини, відомі в сукупності як клітинна оболонка. У прокаріотичних клітинок структури клітинної оболонки змінюються в залежності від типу клітини і організму. Більшість (але не всі) прокаріотичні клітини мають клітинну стінку, але склад цієї клітинної стінки різниться. Всі клітини (прокаріотичні та еукаріотичні) мають плазматичну мембрану (також звану цитоплазматичною мембраною або клітинною мембраною), яка проявляє селективну проникність, дозволяючи деяким молекулам потрапляти або залишати клітину, одночасно обмежуючи прохід інших.
Структура плазматичної мембрани часто описується з точки зору моделі рідкої мозаїки, яка відноситься до здатності мембранних компонентів рухатися по рідини в площині мембрани, а також мозаїчний склад компонентів, які включають різноманітний масив ліпідних і білкових компонентів ( Малюнок\(\PageIndex{10}\)). Структура плазматичної мембрани більшості типів бактерій та еукаріотичних клітин являє собою бішар, що складається в основному з фосфоліпідів, утворених з ефіровими зв'язками та білками. Ці фосфоліпіди та білки мають здатність рухатися латерально всередині площини мембран, а також між двома фосфоліпідними шарами.

Археальні мембрани принципово відрізняються від бактеріальних і еукаріотичних мембран кількома значними способами. По-перше, фосфоліпіди архейної мембрани утворюються з ефірними зв'язками, на відміну від ефірних зв'язків, виявлених в мембранах бактерій або еукаріотичних клітин. По-друге, архейні фосфоліпіди мають розгалужені ланцюги, тоді як у бактеріальних та еукаріотичних клітин прямі ланцюги. Нарешті, хоча деякі архейні мембрани можуть бути сформовані з бішарів, подібних до бактерій та еукаріотів, інші архейні плазматичні мембрани є ліпідними моношарами.
Білки на поверхні клітини важливі для різних функцій, включаючи зв'язок між клітиною та клітиною та зондування умов навколишнього середовища та патогенних факторів вірулентності. Мембранні білки і фосфоліпіди можуть мати вуглеводи (цукру), пов'язані з ними і називаються глікопротеїнами або гліколіпідами відповідно. Ці глікопротеїнові і гліколіпідні комплекси виходять з поверхні клітини, дозволяючи клітці взаємодіяти із зовнішнім середовищем (рис.\(\PageIndex{10}\)). Глікопротеїни і гліколіпіди в плазматичній мембрані можуть значно відрізнятися за хімічним складом серед архей, бактерій та еукаріотів, що дозволяє вченим використовувати їх для характеристики унікальних видів.
Плазмові мембрани з різних типів клітин також містять унікальні фосфоліпіди, які містять жирні кислоти. Як описано в розділі Використання біохімії для ідентифікації мікроорганізмів, профілі аналізу жирних кислот, отриманих фосфоліпідів (PLFA), можуть бути використані для ідентифікації унікальних типів клітин на основі відмінностей в жирних кислотах. Археї, бактерії та еукаріоти мають унікальний профіль PFLA.
Мембранні транспортні механізми
Однією з найважливіших функцій плазматичної мембрани є контроль транспорту молекул всередину і з клітини. Внутрішні умови повинні підтримуватися в певному діапазоні, незважаючи на будь-які зміни зовнішнього середовища. Транспортування речовин через плазматичну мембрану дозволяє клітинам це робити.
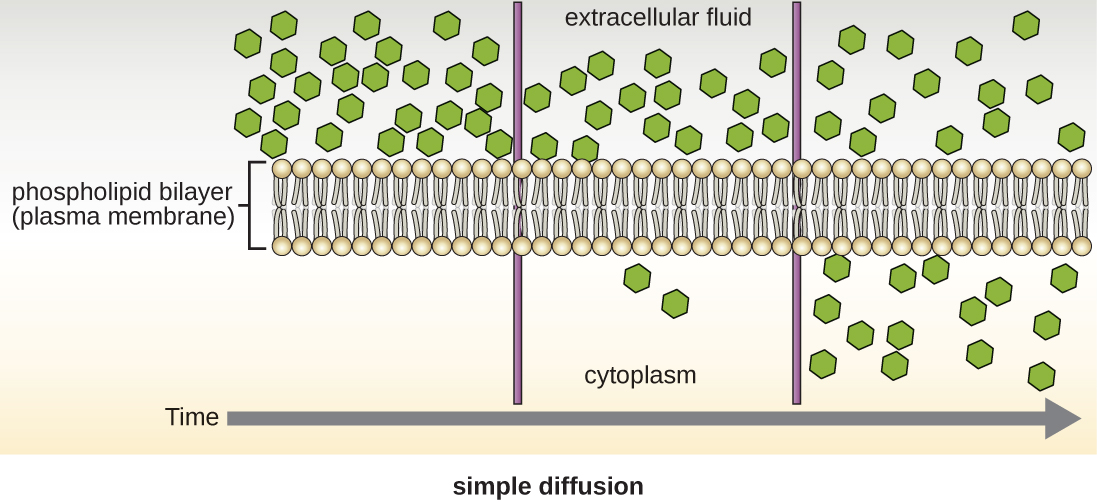
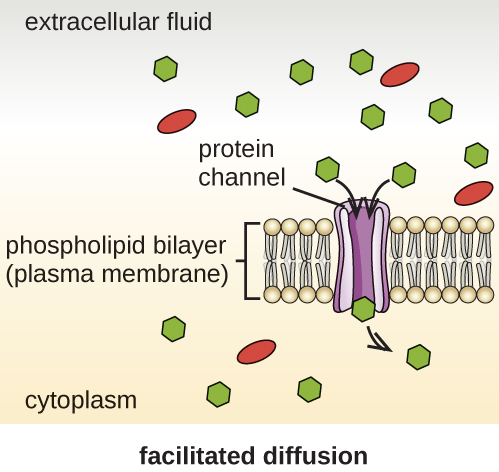
Клітини використовують різні види транспорту через плазматичну мембрану. Наприклад, молекули, що рухаються від більш високої концентрації до більш низької концентрації з градієнтом концентрації, транспортуються простою дифузією, також відомою як пасивний транспорт (рис.\(\PageIndex{11}\)). Деякі дрібні молекули, такі як вуглекислий газ, можуть перетнути мембранний бішар безпосередньо шляхом простої дифузії. Однак заряджені молекули, а також великі молекули потребують допомоги носіїв або каналів в мембрані. Ці структури перетинають молекули через мембрану, процес, відомий як полегшена дифузія (рис.\(\PageIndex{12}\)).
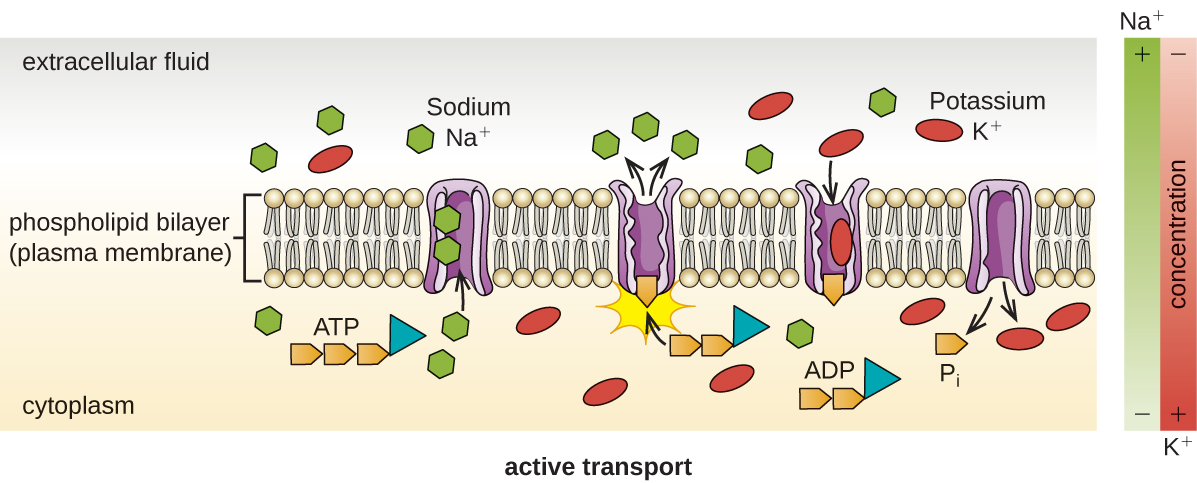
Активний транспорт відбувається, коли клітини рухають молекули через свою мембрану проти градієнтів концентрації (рис.\(\PageIndex{13}\)). Основна відмінність між пасивним і активним транспортом полягає в тому, що активний транспорт вимагає аденозинтрифосфату (АТФ) або інших форм енергії для переміщення молекул «в гору». Тому активні транспортні конструкції часто називають «насосами».
Групова транслокація також транспортує речовини в клітини бактерій. У цьому випадку, коли молекула рухається в клітину проти її градієнта концентрації, вона хімічно модифікується так, що вона не потребує транспортування проти несприятливого градієнта концентрації. Поширеним прикладом цього є бактеріальна система фосфотрансферази, серія носіїв, які фосфорилюють (тобто додає фосфатні іони) глюкозу або інші цукри при попаданні в клітини. Оскільки фосфорилювання цукрів потрібно на ранніх стадіях цукрового обміну, система фосфотрансферази вважається енергонейтральною системою.
Фотосинтетичні мембранні структури
Деякі прокаріотичні клітини, а саме ціанобактерії та фотосинтезуючі бактерії, мають мембранні структури, які дозволяють їм здійснювати фотосинтез. Ці структури складаються з вбудовування плазматичної мембрани, яка охоплює фотосинтетичні пігменти, такі як зелені хлорофіли та бактеріохлорофіли. У ціанобактерій ці мембранні структури називаються тілакоїдами; у фотосинтетичних бактерій їх називають хроматофорами, ламелями або хлоросомами.
Клітинна стінка
Основна функція клітинної стінки полягає в захисті клітини від суворих умов у зовнішньому середовищі. Коли він присутній, є помітні подібності та відмінності між клітинними стінками архей, бактерій та еукаріотів.
Основний компонент клітинних стінок бактерій називається пептидоглікан (або муреїн); він міститься лише в бактеріях. Конструктивно пептидоглікан нагадує шар сітки або тканини (рис.\(\PageIndex{14}\)). Кожен шар складається з довгих ланцюгів, що чергуються молекул N-ацетилглюкозаміну (NAG) і N-ацетилмурамовой кислоти (NAM). Структура довгих ланцюгів має значну двовимірну міцність на розрив завдяки утворенню пептидних містків, які з'єднують NAG і NAM всередині кожного шару пептидоглікану. У грамнегативних бактерій тетрапептидні ланцюги, що відходять від кожної одиниці НАМ, безпосередньо зшиті, тоді як у грампозитивних бактерій ці тетрапептидні ланцюги пов'язані між собою пентагліциновими перехресними мостами. Субодиниці пептидоглікану виготовляються всередині бактеріальної клітини, а потім експортуються та збираються шарами, надаючи клітці її форму.
Оскільки пептидоглікан унікальний для бактерій, багато антибіотичні препарати призначені для втручання в синтез пептидоглікану, послаблюючи клітинну стінку і роблячи клітини бактерій більш сприйнятливими до впливу осмотичного тиску (див. Механізми антибактеріальних препаратів). Крім того, певні клітини імунної системи людини здатні «розпізнавати» бактеріальні збудники шляхом виявлення пептидоглікану на поверхні бактеріальної клітини; ці клітини потім поглинають і руйнують бактеріальну клітину, використовуючи такі ферменти, як лізоцим, який розщеплює і перетравлює пептидоглікан в їх клітинних стінках. (див. Розпізнавання патогенів та фагоцитоз).
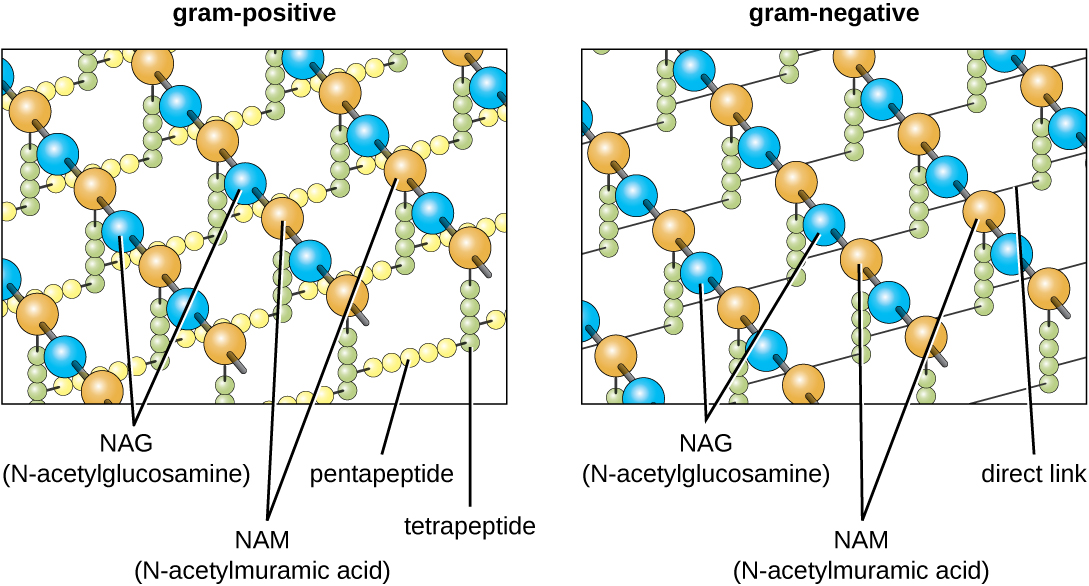
Протокол фарбування за грамом (див. Фарбування мікроскопічних зразків) використовується для диференціації двох поширених типів структур клітинної стінки (рис.\(\PageIndex{15}\)). Грампозитивні клітини мають клітинну стінку, що складається з безлічі шарів пептидоглікану загальною товщиною 30-100 нм. Ці пептидогліканові шари зазвичай вбудовані тейхоєвими кислотами (TAs), вуглеводними ланцюгами, які простягаються через і за межі пептидогліканового шару. Вважається, що 4 TA стабілізує пептидоглікан, збільшуючи його жорсткість. ТА також відіграє роль у здатності патогенних грампозитивних бактерій, таких як Streptococcus, зв'язуватися з певними білками на поверхні клітин господаря, посилюючи їх здатність викликати інфекцію. Крім пептидоглікану та таС, бактерії сімейства Mycobacteriaceae мають зовнішній шар воскових міколівих кислот у своїй клітинній стінці; як описано в Фарбуванні мікроскопічних зразків, ці бактерії називаються кислотостійкими, оскільки кислотостійкі плями повинні використовуватися для проникнення в міколію кислотний шар для цілей мікроскопії (рис.\(\PageIndex{16}\)).

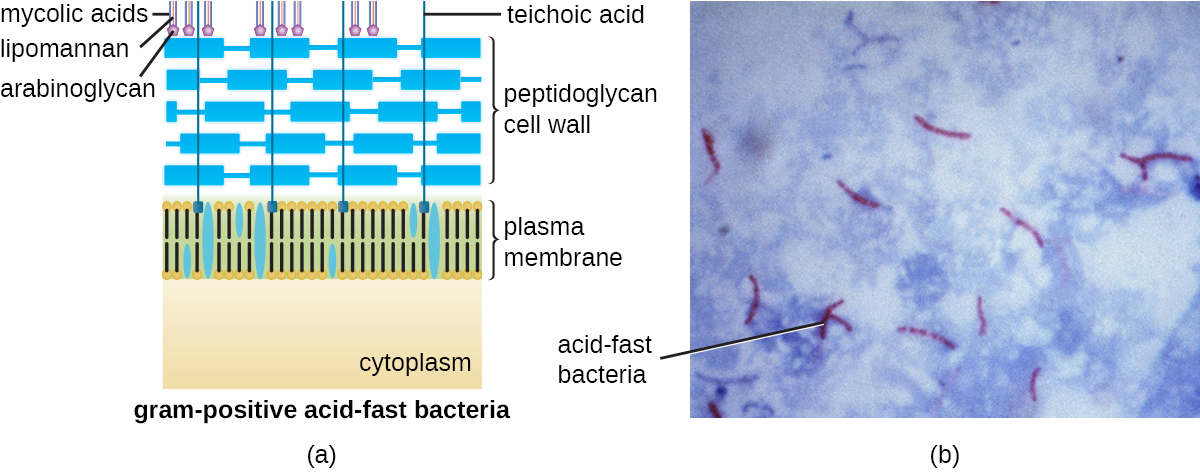
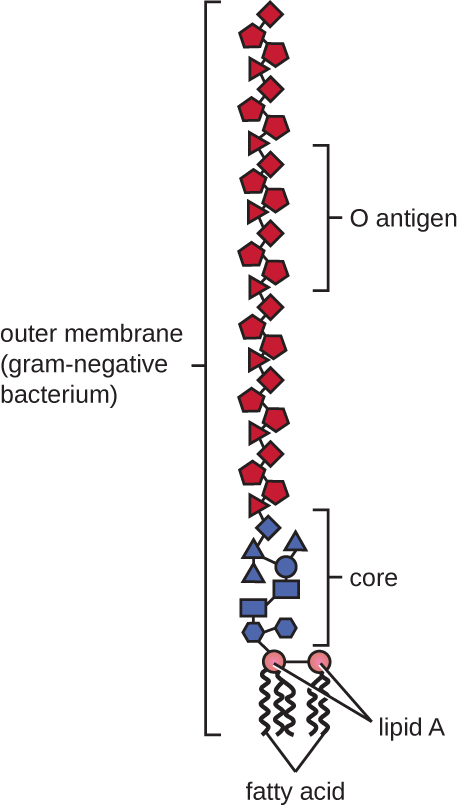
Грамнегативні клітини мають набагато тонший шар пептидоглікану (товщиною не більше 4 нм 6), ніж грампозитивні клітини, а загальна структура їх клітинної оболонки більш складна. У грамнегативних клітині гелеподібний матрикс займає периплазматический простір між клітинною стінкою і плазматичною мембраною, і є другий ліпідний бішар, званий зовнішньою мембраною, який є зовнішнім по відношенню до пептидогліканового шару (рис.\(\PageIndex{15}\)). Ця зовнішня мембрана прикріплена до пептидоглікану ліпопротеїном мурейну. Зовнішня листівка зовнішньої мембрани містить молекулу ліпополісахариду (ЛПС), яка функціонує як ендотоксин при інфекціях, що включають грамнегативні бактерії, сприяючи таким симптомам, як лихоманка, крововилив та септичний шок. Кожна молекула LPS складається з ліпідного А, серцевого полісахариду та бічного ланцюга O, який складається з цукроподібних молекул, які складають зовнішню поверхню ЛПС (рис.\(\PageIndex{17}\)). Склад бічного ланцюга O варіюється між різними видами і штамами бактерій. Частини бокового ланцюга O, звані антигенами, можуть бути виявлені за допомогою серологічних або імунологічних тестів для виявлення конкретних патогенних штамів, таких як Escherichia coli O157: H7, смертельний штам бактерій, який викликає криваву діарею та ниркову недостатність.
Структура клітинної стінки археї відрізняється від структури бактерій кількома значними способами. По-перше, стінки архейних клітин не містять пептидоглікану; натомість вони містять подібний полімер під назвою псевдопептидоглікан (псевдомуреїн), в якому НАМ замінено іншою субодиницею. Інші археї можуть мати шар глікопротеїнів або полісахаридів, який служить клітинною стінкою замість псевдопептидоглікану. Останнє, як і у випадку з деякими бактеріальними видами, є кілька архей, яким, здається, повністю не вистачає клітинних стінок.
Глікокаліці та S-шари
Хоча більшість прокаріотичних клітин мають клітинні стінки, деякі можуть мати додаткові структури оболонки клітин зовні до клітинної стінки, такі як глікокаліці та S-шари. Глікокалікс - це цукрова оболонка, з якої є два важливих типи: капсули і шари слизу. Капсула - це організований шар, розташований зовні клітинної стінки і зазвичай складається з полісахаридів або білків (рис.\(\PageIndex{18}\)). Шар слизу - це менш щільно організований шар, який лише вільно прикріплений до клітинної стінки і може бути легше змивається. Шари слизу можуть складатися з полісахаридів, глікопротеїнів або гліколіпідів.
Глікокаліци дозволяють клітинам прилипати до поверхонь, допомагаючи утворенню біоплівок (колоній мікробів, які утворюються шарами на поверхнях). У природі більшість мікробів живуть у змішаних спільнотах в межах біоплівок, частково тому, що біоплівка забезпечує їм певний рівень захисту. Біоплівки зазвичай утримують воду, як губка, запобігаючи висихання. Вони також захищають клітини від хижацтва і перешкоджають дії антибіотиків і дезінфікуючих засобів. Всі ці властивості вигідні для мікробів, що живуть в біоплівці, але вони представляють проблеми в клінічній обстановці, де метою часто є усунення мікробів.

Здатність виробляти капсулу може сприяти патогенності мікроба (здатність викликати захворювання), оскільки капсула може ускладнити поглинання та вбивання мікроорганізму фагоцитарних клітин (таких як лейкоцити). Наприклад, Streptococcus pneumoniae виробляє капсулу, яка, як відомо, допомагає у патогенності цієї бактерії. Як пояснюється в Фарбування мікроскопічних зразків, капсули важко фарбувати для мікроскопії; Зазвичай використовуються негативні методи фарбування.
S-шар - ще один тип структури оболонки клітин; він складається з суміші структурних білків і глікопротеїнів. У бактерій S-шари знаходяться поза клітинної стінки, але в деяких архей S-шар служить клітинною стінкою. Точна функція S-шарів не зовсім зрозуміла, і їх важко вивчити; але наявні дані свідчать про те, що вони можуть грати різноманітні функції в різних прокаріотичних клітиках, наприклад, допомагати клітині протистояти осмотичному тиску і, для певних патогенів, взаємодіяти з імунним господарем система.
Клінічна спрямованість: Частина 3
Після постановки діагнозу Барбари пневмонія, ПА пише їй рецепт на амоксицилін, зазвичай призначається тип похідного пеніциліну. Більш ніж через тиждень, незважаючи на повний курс за вказівкою, Барбара все ще відчуває слабкість і не повністю одужала, хоча вона все ще здатна пройти через свою повсякденну діяльність. Вона повертається до медичного центру для подальшого візиту.
Багато видів бактерій, грибів та вірусів можуть викликати пневмонію. Амоксицилін націлений на пептидоглікан клітинних стінок бактерій. Оскільки амоксицилін не дозволив симптоми Барбари, ПА робить висновок, що збуднику, ймовірно, не вистачає пептидоглікану, що означає, що збудником може бути вірус, грибок або бактерія, якій не вистачає пептидоглікану. Інша можливість полягає в тому, що збудником є бактерія, що містить пептидоглікан, але має розвинену стійкість до амоксициліну.
Вправа\(\PageIndex{3}\)
- Як ПА може остаточно визначити причину пневмонії Барбари?
- Яку форму лікування повинен призначати ПА, враховуючи, що амоксицилін виявився неефективним?
Ниткоподібні придатки
Багато бактеріальних клітин мають білкові придатки, вбудовані в їх клітинні оболонки, які поширюються назовні, дозволяючи взаємодіяти з навколишнім середовищем. Ці придатки можуть прикріплюватися до інших поверхонь, переносити ДНК або забезпечувати рух. Ниткоподібні придатки включають фімбрії, пілі та джгутики.
Фімбрії і Пілі
Fimbriae і pili структурно схожі, і, оскільки диференціація між ними проблематична, ці терміни часто використовуються взаємозамінно. 7 8 Термін fimbriae зазвичай відноситься до коротких щетиноподібних білків, що виступають з поверхні клітини сотнями. Fimbriae дозволяють клітині прикріплюватися до поверхонь і до інших клітин. Для патогенних бактерій прихильність до клітин господаря важлива для колонізації, інфекційності та вірулентності. Прилипання до поверхонь також має важливе значення при формуванні біоплівки.
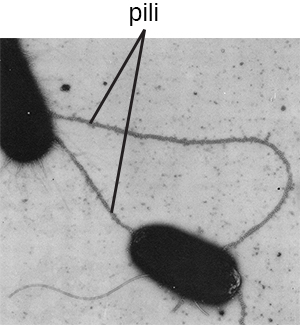
Термін пілі (сингулярний: pilus) зазвичай відноситься до довших, менш численних білкових придатків, які допомагають у прикріпленні до поверхонь (рис.\(\PageIndex{19}\)). Конкретний тип пілуса, який називається F pilus або sex pilus, важливий для передачі ДНК між бактеріальними клітинами, що відбувається між членами одного покоління, коли дві клітини фізично переносять або обмінюються частинами відповідних геномів (див. Як безстатеві прокаріоти досягають генетичного Різноманітність).
Перш ніж структура та функція різних компонентів оболонки клітин бактерій були добре зрозумілі, вчені вже використовували характеристики оболонки клітин для класифікації бактерій. У 1933 році Ребекка Лансфілд запропонувала метод серотипування різних β-гемолітичних штамів видів Streptococcus за допомогою агглютинаційного аналізу, методики з використанням скупчення бактерій для виявлення специфічних клітинних поверхневих антигенів. При цьому Ленсфілд виявив, що одна група S. pyogenes, виявлена в групі А, пов'язана з безліччю захворювань людини. Вона визначила, що різні штами стрептової групи А можна відрізнити один від одного на основі варіацій конкретних білків поверхні клітин, які вона назвала білками M.
Сьогодні на основі протеїнів М ідентифіковано понад 80 різних штамів стрептової групи А. Різні штами стрептококової групи А пов'язані з найрізноманітнішими інфекціями людини, включаючи стрептококовий фарингіт (стрептококовий фарингіт), імпетиго, синдром токсичного шоку, скарлатину, ревматичну лихоманку та некротизуючий фасциит. Білок М є важливим фактором вірулентності стрептової групи А, допомагаючи цим штамам ухилятися від імунної системи. Зміни в білках М змінюють інфекційність певного штаму стрептової групи А.
джгутики
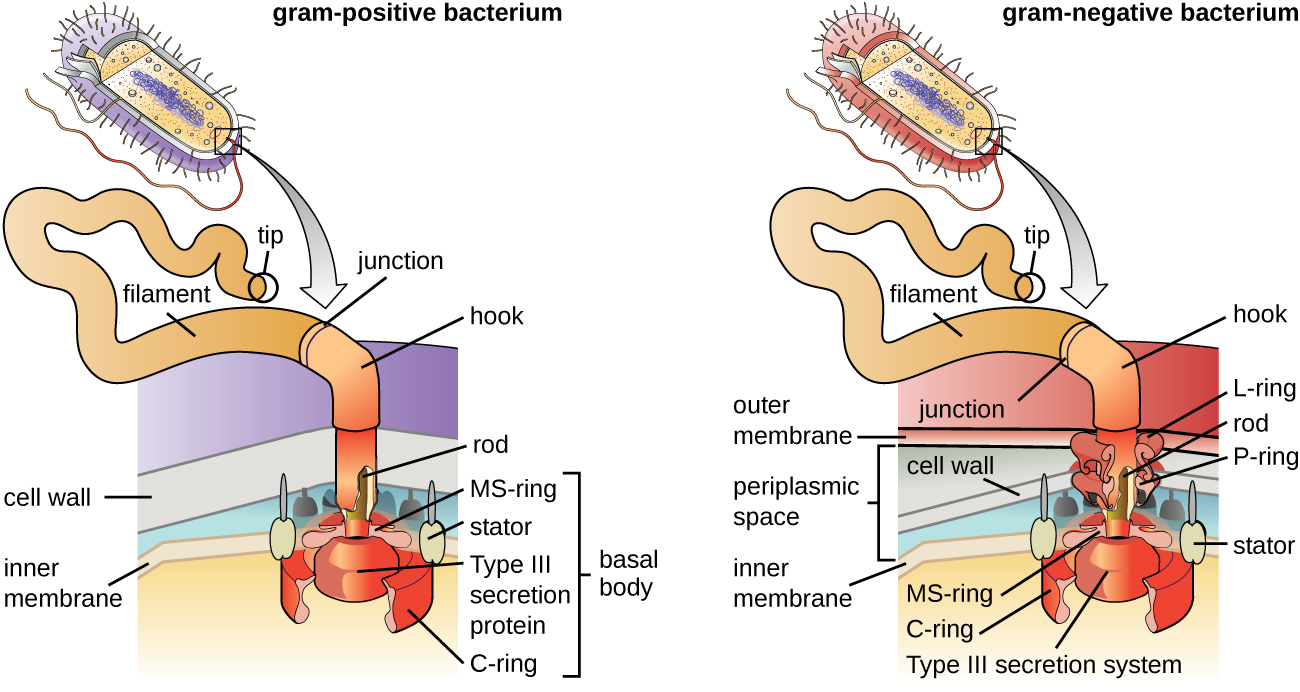
Джгутики - це структури, використовувані клітинами для переміщення у водних середовищах. Бактеріальні джгутики діють подібно пропелерів. Вони являють собою жорсткі спіральні нитки, що складаються з субодиниць білка джгутиків, які виходять назовні від клітини і обертаються в розчині. Базальне тіло є двигуном для джгутика і вбудоване в плазмову мембрану (рис.\(\PageIndex{20}\)). Область гачка з'єднує базальне тіло з ниткою розжарювання. Грампозитивні та грамнегативні бактерії мають різні базальні конфігурації тіла через відмінності в структурі клітинної стінки.
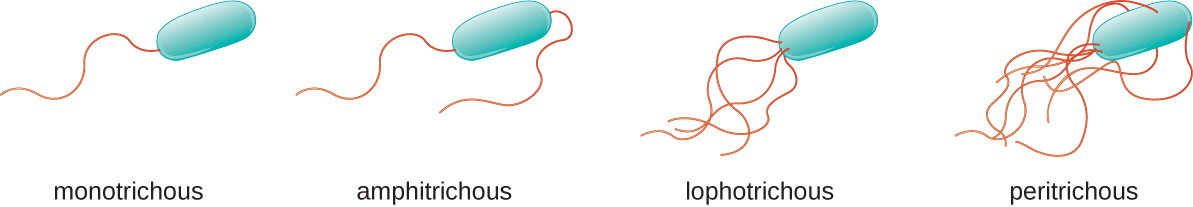
Різні види рухомих бактерій демонструють різне розташування джгутиків (рис.\(\PageIndex{21}\)). Кажуть, що бактерія з сингулярним джгутиком, як правило, розташована на одному кінці клітини (полярної), має монотрихозний джгутик. Прикладом монотрихозно джгутикового бактеріального збудника є Vibrio cholerae, грамнегативна бактерія, яка викликає холеру. Клітини з амфітріческімі джгутиками мають джгутик або пучки джгутиків на кожному кінці. Прикладом може служити Spirillum minor, причина спіриллярної (азіатської) лихоманки укусу щура або содоку. Клітини з лофотриховими джгутиками мають пучок на одному кінці клітини. Грамнегативна паличка Pseudomonas aeruginosa, умовно-патогенний збудник, відомий тим, що викликає багато інфекцій, включаючи інфекції «вуха плавця» та опікові рани, має лофотріхозні джгутики. Джгутики, які покривають всю поверхню бактеріальної клітини, називаються перитріческімі джгутиками. Грамнегативна бактерія E. coli проявляє перитрійное розташування джгутиків.
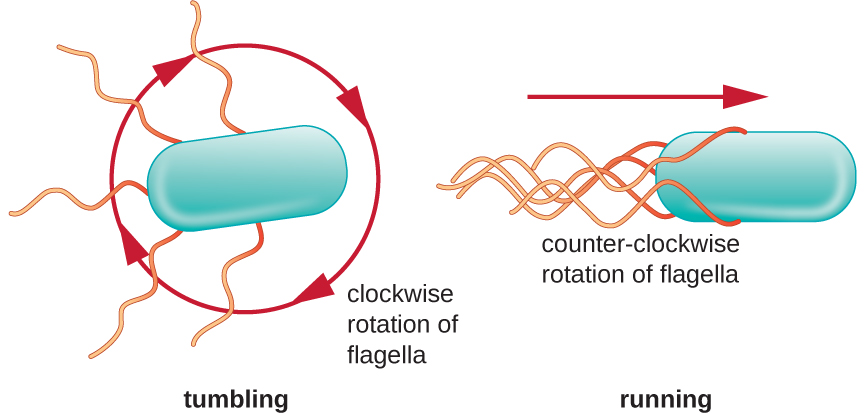
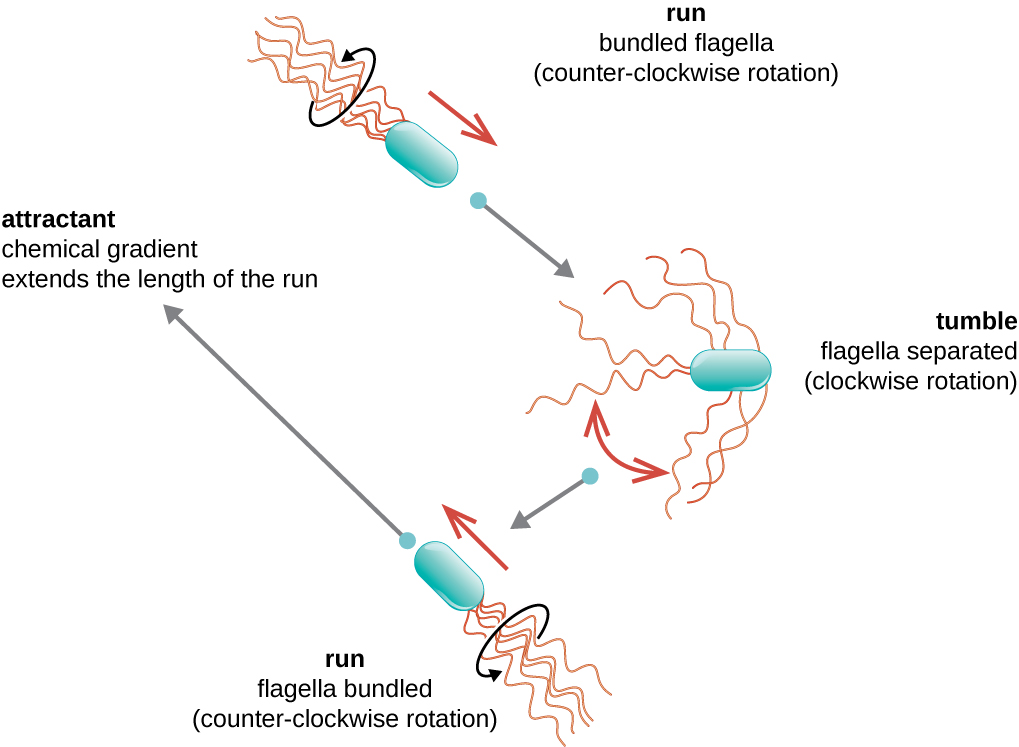
Спрямований рух залежить від конфігурації джгутиків. Бактерії можуть рухатися у відповідь на різноманітні сигнали навколишнього середовища, включаючи світло (фототаксис), магнітні поля (магнітотаксис) за допомогою магнітосом, і, найчастіше, хімічні градієнти (хемотаксис). Цілеспрямований рух в бік хімічного аттрактанта, як джерело їжі, або подалі від репеленту, як отруйна хімічна речовина, досягається збільшенням довжини прогонів і зменшенням довжини тумблів. При бігу джгутики обертаються проти годинникової стрілки, дозволяючи бактеріальної клітці рухатися вперед. У перитрічної бактерії джгутики всі зв'язані між собою дуже обтічним способом (рис.\(\PageIndex{22}\)), що дозволяє ефективно пересуватися. При перекиданні джгутики розкладаються при обертанні за годинниковою стрілкою, створюючи петлевий рух і запобігаючи змістовному руху вперед, але переорієнтуючи клітку в бік атрактанта. Коли аттрактант існує, біги та падіння все ще відбуваються; однак довжина пробіжок довша, тоді як довжина тумбів зменшується, що дозволяє загальний рух у напрямку до вищої концентрації атрактанта. Коли не існує хімічного градієнта, довжини прогонів і перекидів більш рівні, а загальний рух більш випадковий (рис.\(\PageIndex{23}\)).
Вправа\(\PageIndex{4}\)
- Що таке пептидоглікановий шар і чим він відрізняється між грампозитивними і грамнегативними бактеріями?
- Порівняйте і контрастуйте монотрихозні, амфітріческіе, лофотріхові і перитрічні джгутики.
Резюме
- Прокаріотичні клітини відрізняються від еукаріотичних клітин тим, що їх генетичний матеріал міститься в нуклеоїді, а не мембранно-зв'язаному ядрі. Крім того, прокаріотичним клітинам, як правило, не вистачає мембранно-зв'язаних органел.
- Прокаріотичні клітини одного виду зазвичай мають схожу морфологію клітин і клітинне розташування.
- Більшість прокаріотичних клітин мають клітинну стінку, яка допомагає організму підтримувати клітинну морфологію і захищає його від змін осмотичного тиску.
- Поза нуклеоїда прокаріотичні клітини можуть містити екстрахромосомну ДНК в плазмідах.
- Прокаріотичні рибосоми, які знаходяться в цитоплазмі, мають розмір 70S.
- Деякі прокаріотичні клітини мають включення, які зберігають поживні речовини або хімічні речовини для інших цілей.
- Деякі прокаріотичні клітини здатні формувати ендоспори за допомогою споролювання, щоб вижити в стані спокою, коли умови несприятливі. Ендоспори можуть проростати, перетворюючись назад у вегетативні клітини, коли умови покращуються.
- У прокаріотичних клітинок клітинна оболонка включає плазматичну мембрану і зазвичай клітинну стінку.
- Бактеріальні мембрани складаються з фосфоліпідів з інтегральними або периферичними білками. Жирні кислотні компоненти цих фосфоліпідів пов'язані з ефіром і часто використовуються для ідентифікації конкретних типів бактерій. Білки виконують різноманітні функції, включаючи транспорт, зв'язок між клітинами та клітиною та зондування умов навколишнього середовища. Археальні мембрани відрізняються тим, що вони складаються з жирних кислот, які пов'язані ефіром з фосфоліпідами.
- Деякі молекули можуть переміщатися через бактеріальну мембрану шляхом простої дифузії, але більшість великих молекул повинні активно транспортуватися через мембранні структури за допомогою клітинної енергії.
- Прокаріотичні клітинні стінки можуть складатися з пептидоглікану (бактерій) або псевдопептидоглікану (архей).
- Грампозитивні бактеріальні клітини характеризуються товстим пептидоглікановим шаром, тоді як грамнегативні бактеріальні клітини характеризуються тонким пептидоглікановим шаром, оточеним зовнішньою мембраною.
- Деякі прокаріотичні клітини виробляють глікокаліксні покриття, такі як капсули та шари шламу, які допомагають прикріплюватися до поверхонь та/або ухилення від імунної системи господаря.
- Деякі прокаріотичні клітини мають fimbriae або pili, ниткоподібні придатки, які допомагають прикріплюватися до поверхонь. Пілі також використовуються при перенесенні генетичного матеріалу між клітинами.
- Деякі прокаріотичні клітини використовують один або кілька джгутиків для переміщення по воді. Перитріческіе бактерії, які мають численні джгутики, використовують прогони і перекидання, щоб цілеспрямовано рухатися в напрямку хімічного атрактанта.
Виноски
- 1 Р.-Г.М. Чан, В.Ф. Маршалл. «Масштабування властивостей клітин і розмірів органели». Органогенез 6 № 2 (2010) :88—96.
- 2 Ф. Ротфусс, М. Бендер, Р. Конрад. «Виживання та активність бактерій у глибокому, постарілому озерному осадові (Боденське озеро)». Мікробна екологія 33 № 1 (1997) :69—77.
- 3 Р. Сінклер та ін. «Стійкість вибраних агентів категорії А в навколишньому середовищі». Прикладна та екологічна мікробіологія 74 № 3 (2008) :555-563.
- 4 Сілгавий Т.Й., Кане Д., Уокер С.М. «Бактеріальна клітинна оболонка». Перспективи Колд-Спрінг-Харбор в біології 2 № 5 (2010) :a000414.
- 5 Б. Зубер та ін. «Гранульований шар у периплазматичному просторі грампозитивних бактерій та тонких структур Enterococcus gallinarum та Streptococcus gordonii Septa, виявлений шляхом кріо-електронної мікроскопії склоподібних відділів.» Журнал бактеріології 188 № 18 (2006) :6652—6660
- 6 Л.Гана, Чена С.М., Дженсена Г.Дж. «Молекулярна організація грамнегативного пептидоглікану». Праці Національної академії наук Сполучених Штатів Америки 105 № 48 (2008) :18953—18957.
- 7 Дж.А.Гарнетта та співавт. «Структурні уявлення про біогенез та формування біоплівки звичайним пілусом кишкової палички». Праці Національної академії наук Сполучених Штатів Америки 109 № 10 (2012) :3950—3955.
- 8 Т. Профт, Е.Н. Бейкер. «Пілі в грамнегативних і грампозитивних бактеріях - структура, збірка та їх роль у захворюванні». Клітинні та молекулярні науки про життя 66 (2009) :613.