3.4: Унікальні характеристики еукаріотичних клітин
- Page ID
- 4115

Цілі навчання
- Поясніть відмінні характеристики еукаріотичних клітин
- Охарактеризуйте внутрішні та зовнішні структури прокаріотичних клітин з точки зору їх фізичної структури, хімічної структури та функції
- Визначте та опишіть структури та органели, унікальні для еукаріотичних клітин
- Порівняйте та контрастуйте подібні структури, виявлені в прокаріотичних та еукаріотичних клітиках
Еукаріотичні організми включають найпростіших, водоростей, грибів, рослин і тварин. Деякі еукаріотичні клітини є самостійними одноклітинними мікроорганізмами, тоді як інші входять до складу багатоклітинних організмів. Клітини еукаріотичних організмів мають кілька відмінних характеристик. Перш за все, еукаріотичні клітини визначаються наявністю ядра, оточеного складною ядерною мембраною. Також еукаріотичні клітини характеризуються наявністю в цитоплазмі мембрано-зв'язаних органел. Органели, такі як мітохондрії, ендоплазматичний ретикулум (ER), апарат Гольджі, лізосоми та пероксисоми утримуються на місці цитоскелетом, внутрішньою мережею, яка підтримує транспорт внутрішньоклітинних компонентів і допомагає підтримувати форму клітин (рис.\(\PageIndex{1}\)). Геном еукаріотичних клітин упакований у множинні стрижнеподібні хромосоми на відміну від однієї хромосоми круглої форми, яка характеризує більшість прокаріотичних клітин. Таблиця\(\PageIndex{1}\) порівнює характеристики еукаріотичних клітинних структур з бактеріями та архей.

| клітинна структура | Прокаріоти | Еукаріот | |
|---|---|---|---|
| Бактерії | Архея | ||
| Розмір | ~ 0,5—1 мкм | ~ 0,5—1 мкм | ~5-20 мкм |
| Співвідношення площі поверхні до об'єму | Високі | Високі | Низький |
| Ядро | Ні | Ні | Так |
| характеристики генома |
|
|
|
| Розподіл клітин | двійковий поділ | двійковий поділ | Мітоз, мейоз |
| Мембранний ліпідний склад |
|
|
|
| Склад клітинної стінки |
|
|
|
| Рухливі структури | Жорсткі спіральні джгутики, що складаються з джгутиків | Жорсткі спіральні джгутики, що складаються з архейних джгутиків | Гнучкі джгутики і вії, що складаються з мікротрубочок |
| Зв'язані мембранами органели | Ні | Ні | Так |
| Ендомембранна система | Ні | Ні | Так (ER, Гольджі, лізосоми) |
| Рибосоми | 70-ТІ | 70-ТІ |
|
Морфологія клітин
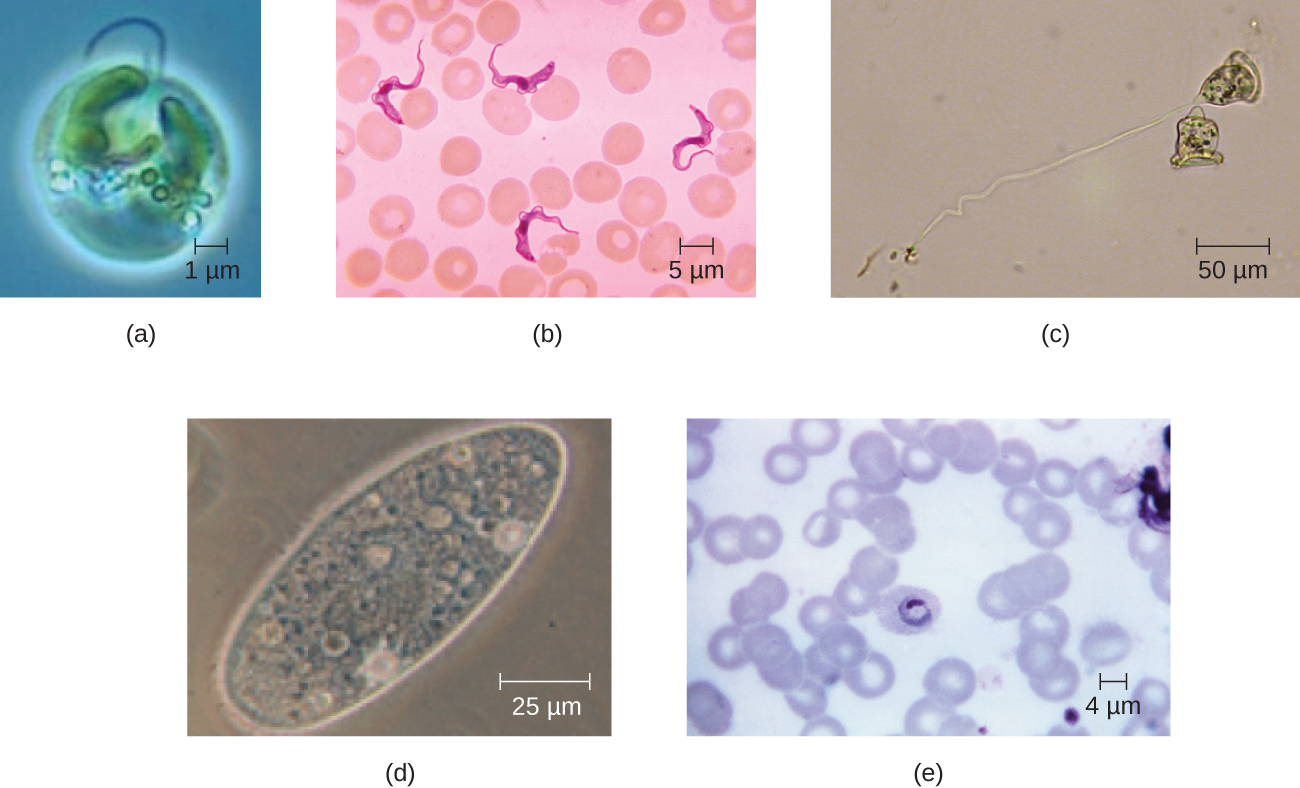
Еукаріотичні клітини демонструють велику різноманітність різних морфологій клітин. Можливі форми включають сфероїдні, яйцеподібні, кубоїдальні, циліндричні, плоскі, лінзоподібні, веретеноподібні, дискоїдні, серповидні, кільцеві зірчасті та багатокутні (рис.\(\PageIndex{2}\)). Деякі еукаріотичні клітини мають неправильну форму, а деякі здатні змінювати форму. На форму конкретного типу еукаріотичної клітини можуть впливати такі фактори, як її первинна функція, організація її цитоскелета, в'язкість її цитоплазми, жорсткість її клітинної мембрани або клітинної стінки (якщо вона є), а також фізичний тиск, що чиниться на неї навколишнім середовищем і/або прилеглі клітини.
Вправа\(\PageIndex{1}\)
Визначте дві відмінності між еукаріотичною та прокаріотичною клітиною.
Ядро
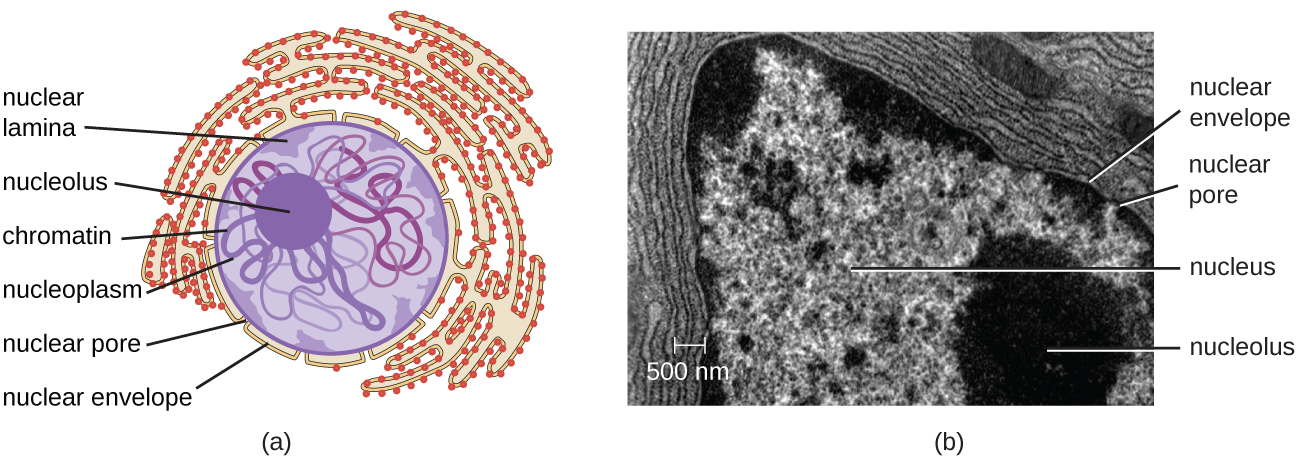
На відміну від прокаріотичних клітин, в яких ДНК нещільно міститься в нуклеоїдної області, еукаріотичні клітини мають ядро, яке оточене складною ядерною мембраною, в якій знаходиться геном ДНК (рис.\(\PageIndex{3}\)). Містивши ДНК клітини, ядро в кінцевому підсумку контролює всю діяльність клітини, а також відіграє важливу роль у розмноженні та спадковості. Еукаріотичні клітини зазвичай мають свою ДНК, організовану в кілька лінійних хромосом. ДНК всередині ядра високоорганізована і конденсується, щоб поміститися всередині ядра, що досягається шляхом обгортання ДНК навколо білків, званих гістонами.
Хоча більшість еукаріотичних клітин мають лише одне ядро, винятки існують. Наприклад, найпростіші роду Paramecium зазвичай мають два повних ядра: невелике ядро, яке використовується для розмноження (мікроядро) і велике ядро, яке направляє клітинний метаболізм (макронуклеус). Крім того, деякі гриби тимчасово утворюють клітини з двома ядрами, званими гетерокаріотичними клітинами, під час статевого розмноження. Клітини, ядра яких діляться, а цитоплазма яких немає, називаються коеноцитами.
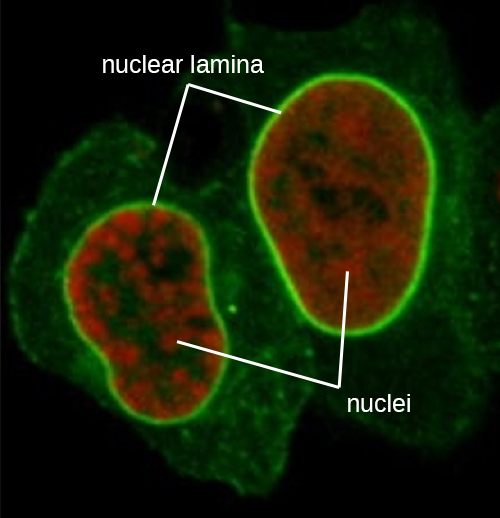
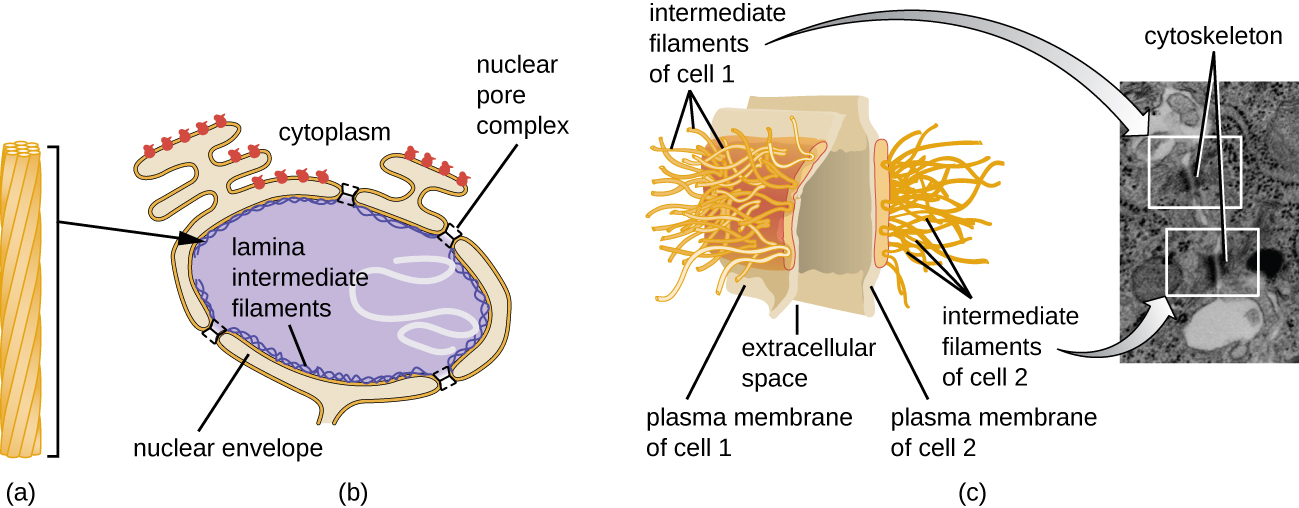
Ядро пов'язане складною ядерною мембраною, яку часто називають ядерною оболонкою, яка складається з двох різних ліпідних бішарів, які є суміжними один з одним (рис.\(\PageIndex{4}\)). Незважаючи на ці зв'язки між внутрішньою і зовнішньою мембранами, кожна мембрана містить унікальні ліпіди і білки на її внутрішній і зовнішній поверхнях. Ядерна оболонка містить ядерні пори, які представляють собою великі, розеткоподібні білкові комплекси, які контролюють рух матеріалів всередину і з ядра. Загальна форма ядра визначається ядерною пластинкою, сіткою проміжних ниток, знайдених якраз всередині мембран ядерної оболонки. Зовні ядра додаткові проміжні нитки утворюють більш пухку сітку і служать для закріплення ядра в положенні всередині клітини.
Ядрыця
Ядро - це щільна область всередині ядра, де відбувається біосинтез рибосомальної РНК (рРНК). Крім того, ядерце також є місцем, де починається збірка рибосом. Прерибосомальні комплекси збираються з рРНК і білків в ядерці; вони потім транспортуються в цитоплазму, де завершується збірка рибосом (рис.\(\PageIndex{5}\)).
Рибосоми
Рибосоми, знайдені в еукаріотичних органелах, таких як мітохондрії або хлоропласти, мають 70-ті рибосоми - такий же розмір, як і прокаріотичні рибосоми. Однак неорганелла-асоційовані рибосоми в еукаріотичних клітині - це рибосоми 80S, що складаються з малої субодиниці 40S та великої субодиниці 60S. За розміром і складом це робить їх відмінними від рибосом прокаріотичних клітин.
Два типи неорганельно-асоційованих еукаріотичних рибосом визначаються їх розташуванням у клітці: вільні рибосоми та мембранозв'язані рибосоми. Вільні рибосоми знаходяться в цитоплазмі і служать для синтезу водорозчинних білків; мембранозв'язані рибосоми виявляються прикріпленими до грубої ендоплазматичної сітки і роблять білки для введення в клітинну мембрану або білки, призначені для експорту з клітини.
Відмінності між еукаріотичними та прокаріотичними рибосомами є клінічно актуальними, оскільки певні антибіотичні препарати призначені для націлювання на ту чи іншу. Наприклад, циклогексимід спрямований на еукаріотичну дію, тоді як хлорамфенікол спрямований на прокаріотичні рибосоми. 1 Оскільки клітини людини є еукаріотичними, їм, як правило, не шкодять антибіотики, які руйнують прокаріотичні рибосоми в бактеріях. Однак іноді можуть виникнути негативні побічні ефекти через те, що мітохондрії в клітині людини містять прокаріотичні рибосоми.
Ендомембранна система
Ендомембранна система, унікальна для еукаріотичних клітин, являє собою ряд мембранозних канальців, мішків і сплющених дисків, які синтезують багато клітинних компонентів і переміщують матеріали навколо клітини (рис.\(\PageIndex{6}\)). Через більший розмір клітин еукаріотичним клітинам потрібна ця система для транспортування матеріалів, які не можуть бути дисперговані лише дифузією. Ендомембранна система містить кілька органел і зв'язків між ними, включаючи ендоплазматичний ретикулум, апарат Гольджі, лізосоми та везикули.
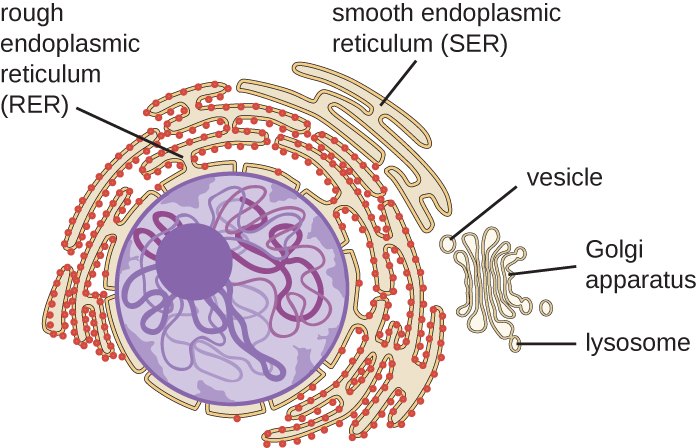
Ендоплазматичний ретикулум
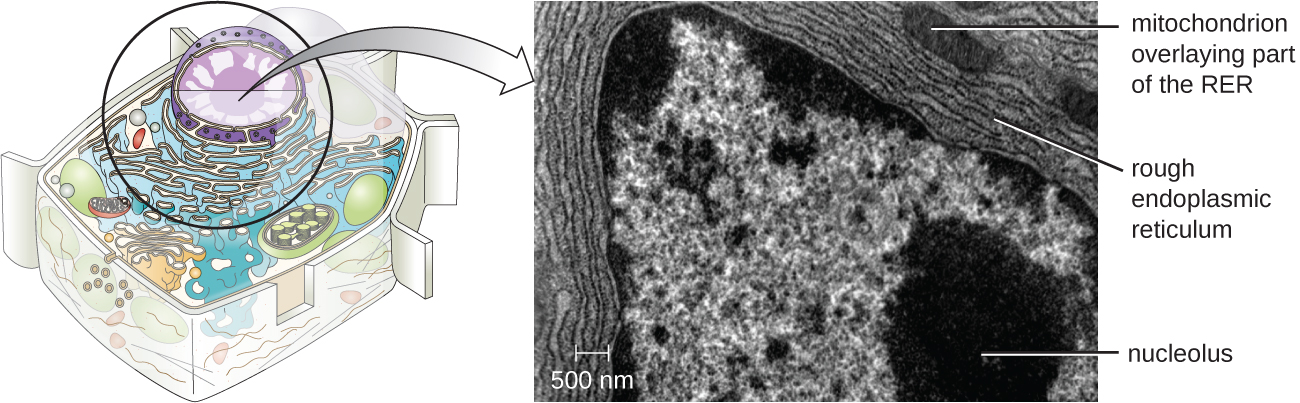
Ендоплазматичний ретикулум (ЕР) являє собою з'єднаний між собою масив канальців і цистерн (сплющених мішечків) з єдиним ліпідним бішаром (рис.\(\PageIndex{7}\)). Прогалини всередині цистерн називаються просвітом ЕР. Існує два типи ER, шорсткий ендоплазматичний ретикулум (RER) та гладкий ендоплазматичний ретикулум (SER). Ці два різних типи ER є сайтами для синтезу чітко різних типів молекул. РЕР шипований рибосомами, пов'язаними на цитоплазматичної стороні мембрани. Ці рибосоми роблять білки, призначені для плазматичної мембрани (рис.\(\PageIndex{}\)). Після синтезу ці білки вставляються в мембрану РЕР. Маленькі мішечки RER, що містять ці знову синтезовані білки, потім відходять як транспортні бульбашки і рухаються або до апарату Гольджі для подальшої обробки, безпосередньо до плазматичної мембрани, до мембрани іншої органели, або з клітини. Транспортні везикули - це одноліпідні, двошарові, мембранозні сфери з порожнистими інтер'єрами, які несуть молекули. СЕР не має рибосом і, отже, виглядає «гладкою». Він бере участь у біосинтезі ліпідів, вуглеводному обміні та детоксикації токсичних сполук всередині клітини.
Апарат Гольджі
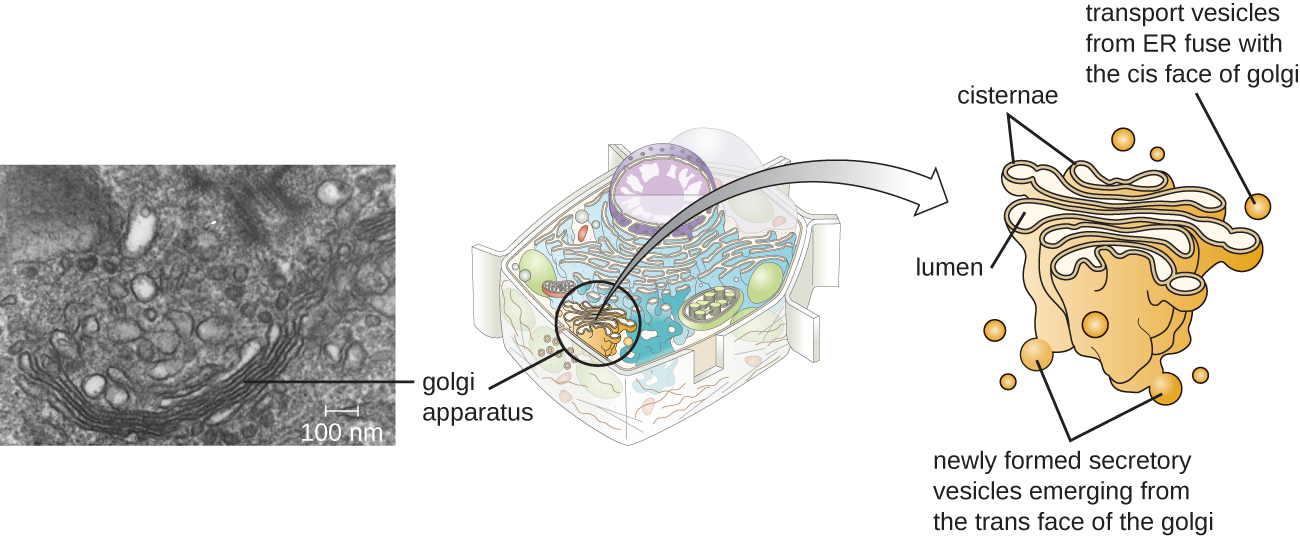
Апарат Гольджі був відкритий в ендомембранній системі в 1898 році італійським вченим Камілло Гольджі (1843—1926), який розробив нову техніку фарбування, яка показала складені мембранні структури всередині клітин плазмодію, збудника малярії. Апарат Гольджі складається з серії мембранозних дисків, які називаються dictyosomes, кожен з яких має один ліпідний бішар, які складені разом (рис.\(\PageIndex{8}\)).
Ферменти в апараті Гольджі модифікують ліпіди і білки, що транспортуються з ЕР в Гольджі, часто додаючи до них вуглеводні компоненти, виробляючи гліколіпіди, глікопротеїни або протеоглікани. Гліколіпіди і глікопротеїни часто вводяться в плазматичну мембрану і важливі для розпізнавання сигналу іншими клітинами або інфекційними частинками. Різні типи клітин можна відрізнити один від одного за будовою і розташуванням гліколіпідів і глікопротеїнів, що містяться в їх плазматичних мембранах. Ці гліколіпіди та глікопротеїни зазвичай також служать рецепторами поверхні клітин.
Транспортуйте бульбашки, що виходять з запобіжника ER з апаратом Гольджі на його прийомі, або цис, обличчі. Білки обробляються в апараті Гольджі, а потім додаткові транспортні везикули, що містять модифіковані білки і ліпіди, відщипують від апарату Гольджі на його вихідному, або транс, обличчі. Ці вихідні бульбашки переміщаються і зливаються з плазматичною мембраною або мембраною інших органел.
Екзоцитоз - це процес, за допомогою якого секреторні бульбашки (кулясті мембранозні мішечки) виділяють свій вміст до зовнішньої сторони клітини (рис.\(\PageIndex{8}\)). Всі клітини мають складові секреторні шляхи, в яких секреторні бульбашки транспортують розчинні білки, які вивільняються з клітини постійно (конституційно). Деякі спеціалізовані клітини також мають регульовані секреторні шляхи, які використовуються для зберігання розчинних білків у секреторних бульбашках. Регульована секреція включає речовини, які виділяються лише у відповідь на певні події або сигнали. Наприклад, певні клітини імунної системи людини (наприклад, огрядні клітини) виділяють гістамін у відповідь на присутність сторонніх предметів або патогенів в організмі. Гістамін - це з'єднання, яке запускає різні механізми, що використовуються імунною системою для усунення хвороботворних мікроорганізмів.
Лізосоми
У 1960-х роках бельгійський вчений Крістіан де Дюв (1917—2013) відкрив лізосоми, пов'язані мембранами органели ендомембранної системи, які містять травні ферменти. Деякі типи еукаріотичних клітин використовують лізосоми для розщеплення різних частинок, таких як їжа, пошкоджені органели або клітинні сміття, мікроорганізми або імунні комплекси. Компартменталізація травних ферментів всередині лізосоми дозволяє клітині ефективно перетравлювати речовину, не завдаючи шкоди цитоплазматичним компонентам клітини.
Вправа\(\PageIndex{2}\)
Назвіть компоненти ендомембранної системи і опишіть функцію кожного компонента.
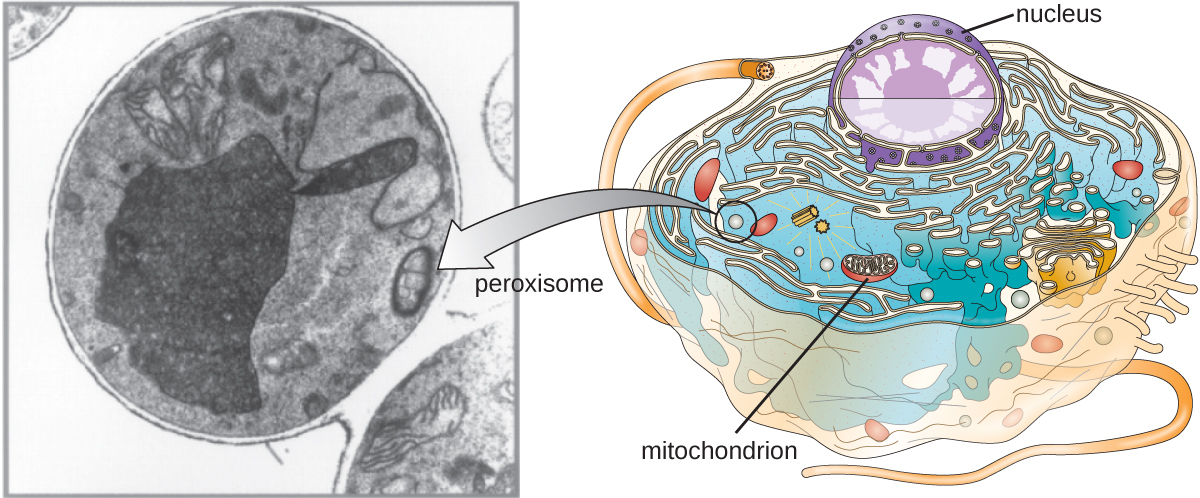
пероксисоми
Крістіану де Дюве також приписують відкриття пероксисом, мембранно-зв'язаних органел, які не входять до складу ендомембранної системи (рис.\(\PageIndex{9}\)). Пероксисоми утворюються незалежно в цитоплазмі від синтезу пероксинових білків вільними рибосомами і включення цих пероксинових білків в існуючі пероксисоми. Зростаючі пероксисоми потім ділять за процесом, подібним до двійкового ділення.
Пероксисоми були вперше названі за їх здатність виробляти перекис водню, високореактивну молекулу, яка допомагає розщеплювати молекули, такі як сечова кислота, амінокислоти та жирні кислоти. Пероксисоми також володіють ферментом каталазою, який може деградувати перекис водню. Поряд з СЕР пероксисоми також відіграють роль у біосинтезі ліпідів. Як і лізосоми, компартменталізація цих деградаційних молекул всередині органели допомагає захистити цитоплазматичний вміст від небажаних пошкоджень.
Пероксисоми певних організмів спеціалізуються для задоволення їх конкретних функціональних потреб. Наприклад, гліоксисоми - це модифіковані пероксисоми дріжджів і рослинних клітин, які виконують кілька метаболічних функцій, включаючи вироблення молекул цукру. Аналогічно глікосоми - це модифіковані пероксисоми, виготовлені певними трипаносомами, патогенними найпростішими, які викликають хворобу Шагаса та африканську сонну хворобу.
Цитоскелет
Еукаріотичні клітини мають внутрішній цитоскелет, складений з мікрофіламентів, проміжних ниток і мікротрубочок. Ця матриця волокон і трубок забезпечує структурну підтримку, а також мережу, по якій матеріали можуть транспортуватися всередині клітини і на якій можуть бути закріплені органели (рис.\(\PageIndex{10}\)). Наприклад, процес екзоцитозу передбачає переміщення бульбашки по цитоскелетної мережі до плазматичної мембрани, де він може вивільняти свій вміст.
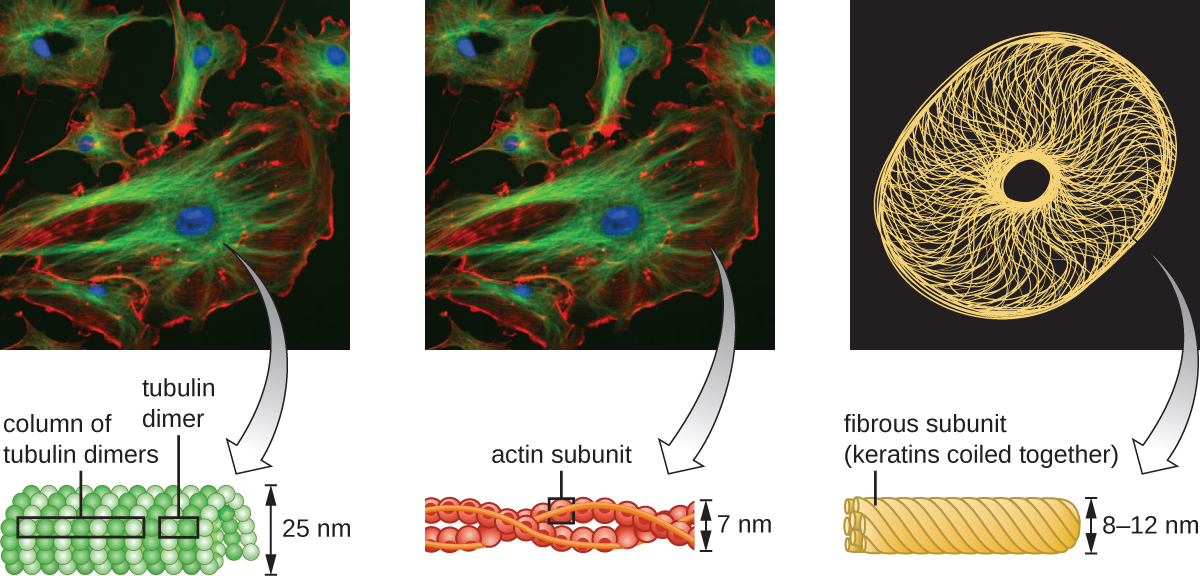
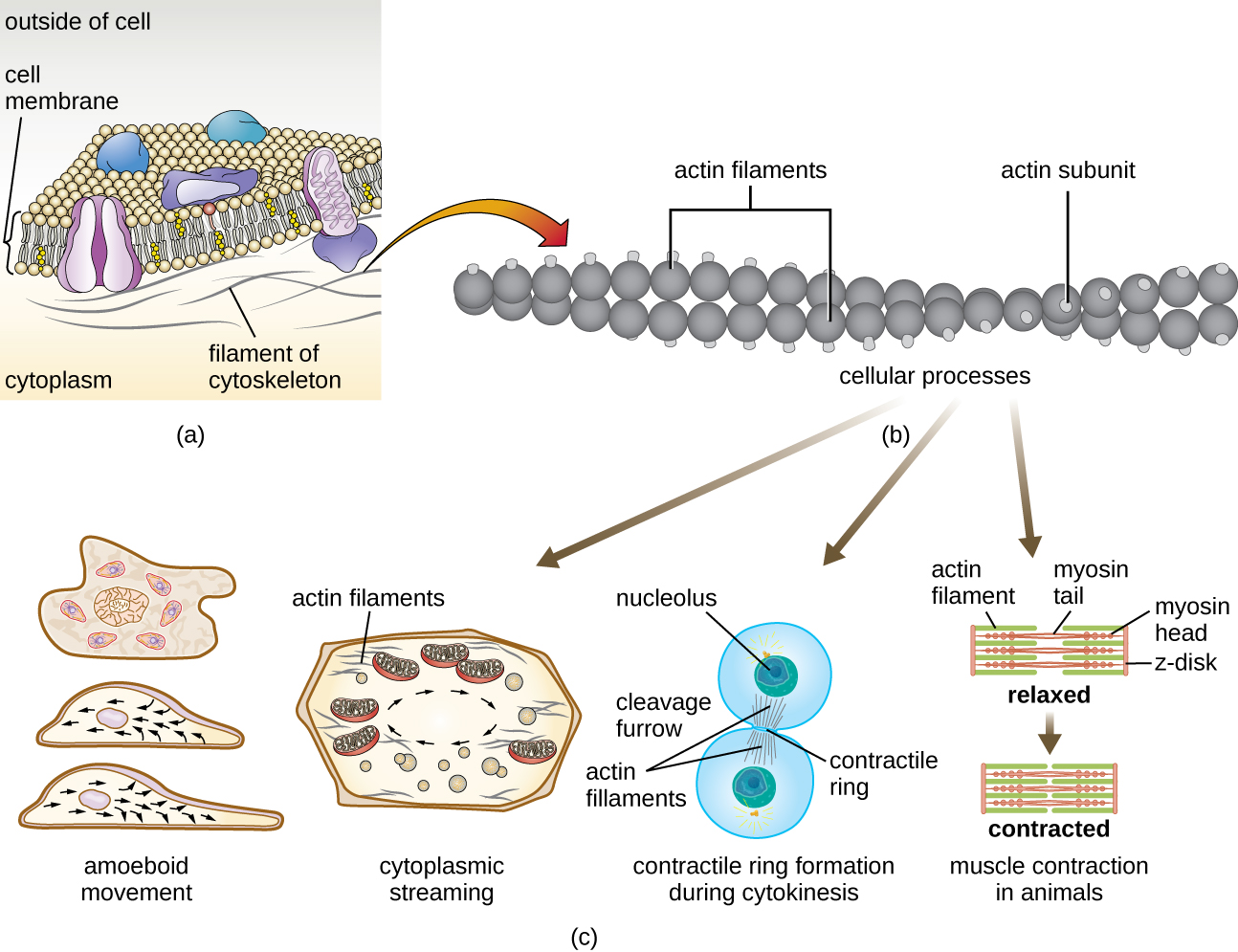
Мікронитки складаються з двох переплетених між собою ниток актину, кожна з яких складається з актинових мономерів, що утворюють ниткоподібні кабелі діаметром 6 нм (рис.\(\PageIndex{11}\)). Актинові нитки працюють разом з руховими білками, такими як міозин, щоб впливати на скорочення м'язів у тварин або амебоїдний рух деяких еукаріотичних мікробів. У амебоїдних організмах актин можна знайти в двох формах: більш жорстка, полімеризована, гелева форма і більш рідка, неполімеризованная розчинна форма. Актин в гелевій формі створює стабільність в ектоплазмі, гелеподібної області цитоплазми якраз всередині плазматичної мембрани амебоїдних найпростіших.
Тимчасові розширення цитоплазматичної мембрани, звані псевдоподіями (що означає «помилкові стопи»), виробляються через прямий потік розчинних актинових ниток в псевдоподію з подальшим циклічним циклічним циклом актинових ниток, що призводить до рухливості клітин. Як тільки цитоплазма поширюється назовні, утворюючи псевдоподіум, залишилася цитоплазма стікає вгору, щоб приєднатися до переднього краю, тим самим створюючи рух вперед. Крім амебоїдного руху, мікрофіламенти також беруть участь у безлічі інших процесів в еукаріотичних клітині, включаючи цитоплазматичне потокове (рух або циркуляція цитоплазми всередині клітини), формування розщеплення борозни при поділі клітин і рух м'язів у тварин (рис.\(\PageIndex{11}\)). Ці функції є результатом динамічного характеру мікрофіламентів, які можуть відносно легко полімеризуватися і деполімеризуватися у відповідь на клітинні сигнали, і їх взаємодії з молекулярними двигунами в різних типах еукаріотичних клітин.
Проміжні нитки (рис.\(\PageIndex{12}\)) - це різноманітна група цитоскелетних ниток, які діють як кабелі всередині клітини. Їх називають «проміжними», оскільки їх 10-нм діаметр товщі, ніж у актину, але тонший, ніж у мікротрубочок. 3 Вони складаються з декількох ниток полімеризованих субодиниць, які, в свою чергу, складаються з найрізноманітніших мономерів. Проміжні нитки мають тенденцію бути більш постійними в клітці і зберігають положення ядра. Вони також утворюють ядерну пластинку (підкладку або шар) безпосередньо всередині ядерної оболонки. Крім того, проміжні нитки відіграють певну роль у закріпленні клітин разом у тканині тварин. Проміжний білок нитки десмін міститься в десмосомах, білкових структурах, які з'єднують м'язові клітини разом і допомагають їм протистояти зовнішнім фізичним силам. Проміжний кератин білка нитки - це структурний білок, який міститься у волоссі, шкірі та нігтях.
Мікротрубочки (рис.\(\PageIndex{13}\)) - це третій тип цитоскелетного волокна, що складається з димерів тубуліну (α тубуліну і β тубуліну). Вони утворюють порожнисті трубки діаметром 23 нм, які використовуються як прогони всередині цитоскелета. 4 Як і мікрофіламенти, мікротрубочки динамічні і мають здатність швидко збирати і розбирати. Мікротрубочки також працюють з руховими білками (такими як динеїн та кінезин), щоб переміщати органели та везикули навколо цитоплазми. Крім того, мікротрубочки є основними компонентами еукаріотичних джгутиків і вій, що складають як нитку, так і базальні компоненти тіла (рис.\(\PageIndex{20}\)).
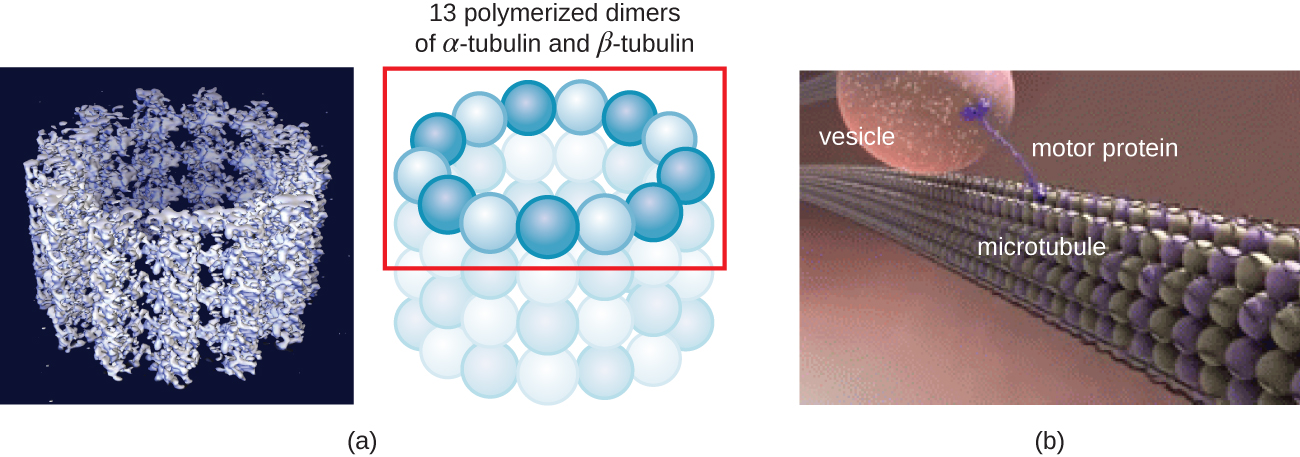
Крім того, мікротрубочки беруть участь в поділі клітин, утворюючи мітотичний веретено, який служить для відділення хромосом під час мітозу і мейозу. Мітотичний веретено виробляється двома центросомами, які по суті є центрами, що організують мікротрубочки, на протилежних кінцях клітини. Кожна центросома складається з пари центриолей, розташованих під прямим кутом один до одного, і кожна центриоль являє собою масив з дев'яти паралельних мікротрубочок, розташованих в трійках (рис.\(\PageIndex{14}\)).
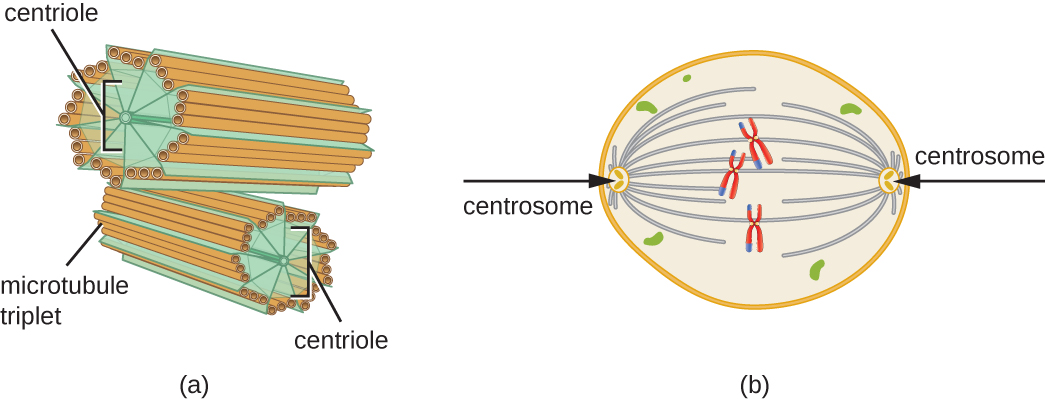
Вправа\(\PageIndex{3}\)
Порівняйте та порівняйте три типи цитоскелетних структур, описаних у цьому розділі.
Мітохондрії
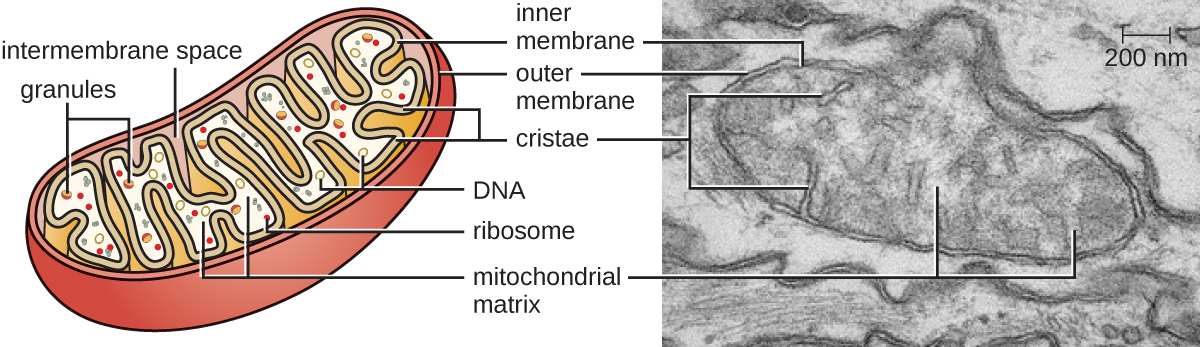
Великі складні органели, в яких відбувається аеробне клітинне дихання в клітинах-еукаріотів, називаються мітохондріями (рис.\(\PageIndex{15}\)). Термін «мітохондріон» був вперше придуманий німецьким мікробіологом Карлом Бендою в 1898 році і пізніше був пов'язаний з процесом дихання Отто Варбургом в 1913 році. Вчені протягом 1960-х років виявили, що мітохондрії мають власний геном і рибосоми 70-х років. Геном мітохондрій виявився бактеріальним, коли він був секвенований у 1976 році. Ці висновки в кінцевому підсумку підтримали ендосимбіотичну теорію, запропоновану Лінн Маргуліс, яка стверджує, що мітохондрії спочатку виникли через ендосимбіотичну подію, в якій бактерія, здатна до аеробного клітинного дихання, була захоплена фагоцитозом у клітину-господаря і залишалася життєздатною внутрішньоклітинною складова.
Кожен мітохондріон має дві ліпідні мембрани. Зовнішня мембрана є залишком мембранних структур вихідної клітини господаря. Внутрішня мембрана була отримана з бактеріальної плазматичної мембрани. Ланцюг транспорту електронів для аеробного дихання використовує інтегральні білки, вбудовані у внутрішню мембрану. Мітохондріальний матрикс, відповідний розташуванню цитоплазми вихідної бактерії, є поточним місцем розташування багатьох метаболічних ферментів. Він також містить мітохондріальну ДНК і рибосоми 70S. Інвагінації внутрішньої мембрани, звані кристами, еволюціонували для збільшення площі поверхні для розташування біохімічних реакцій. Складні візерунки кристи відрізняються між різними типами еукаріотичних клітин і використовуються для відрізнення різних еукаріотичних організмів один від одного.
хлоропласти
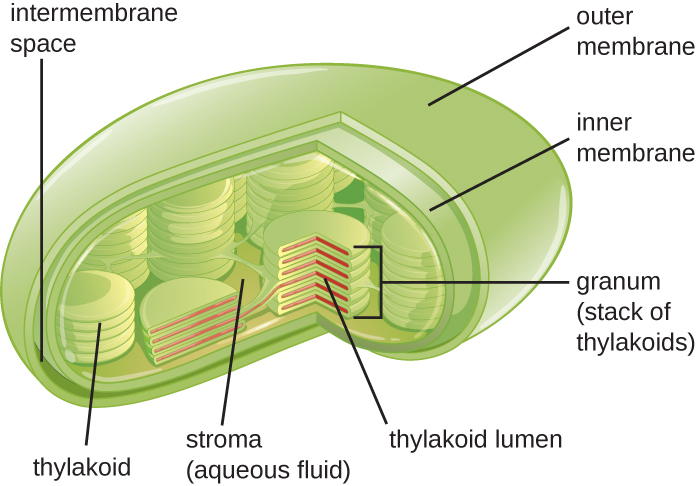
Рослинні клітини і клітини водоростей містять хлоропласти - органели, в яких відбувається фотосинтез (рис.\(\PageIndex{16}\)). Всі хлоропласти мають принаймні три мембранні системи: зовнішню мембрану, внутрішню мембрану та тилакоїдну мембранну систему. Усередині зовнішньої та внутрішньої мембран знаходиться строма хлоропласту, гелеподібна рідина, яка становить значну частину обсягу хлоропласту, і в якій плаває тилакоїдна система. Тилакоїдна система являє собою високодинамічну колекцію складчастих мембранних мішків. Саме там знаходиться зелений фотосинтетичний пігмент хлорофіл і відбуваються світлові реакції фотосинтезу. У більшості рослинних хлоропластів тилакоїди розташовані в стеках, званих грана (сингулярний: granum), тоді як в деяких водорослевих хлоропластах тилакоїди вільно плавають.
Інші органели, схожі з мітохондріями, виникли у інших видів еукаріотів, але їх ролі відрізняються. Гідрогеносоми знаходяться в деяких анаеробних еукаріотів і служать місцем виробництва анаеробного водню. Гідрогеносом зазвичай не вистачає власної ДНК та рибосом. Кінетопласти - це різновид мітохондрій, виявлених у деяких еукаріотичних збудників. У цих організмів кожна клітина має єдиний, довгий, розгалужений мітохондріон, в якому ДНК кінетопласта, організована у вигляді декількох кругових шматочків ДНК, виявляється зосередженою на одному полюсі клітини.
Органели, пов'язані з мітохондріями, у найпростіших паразитів
Багато найпростіших, в тому числі кілька найпростішихпаразитів, що викликають інфекції у людини, можна визначити за їх незвичайним зовнішнім виглядом. Відмінними рисами можуть бути складні морфології клітин, наявність унікальних органел або відсутність загальних органел. Найпростіші паразити лямблії лямблії та Trichomonas vaginalis - два приклади.
Г. лямблії, часта причина діареї у людини і багатьох інших тварин, - анаеробний паразит, який володіє двома ядрами і декількома джгутиками. Його апарат Гольджі і ендоплазматичний ретикулум сильно скорочені, а мітохондрії йому повністю відсутні. Однак у нього є органели, відомі як мітосоми, органели з подвійною мембраною, які, як видається, сильно зменшені мітохондрії. Це змусило вчених вважати, що предки Г. лямблії колись володіли мітохондріями, які еволюціонували, щоб стати мітосомами. T. vaginalis, який викликає венеричний інфекційний вагініт, є ще одним найпростішим паразитам, якому не вистачає звичайних мітохондрій. Натомість він має гідрогеносоми, пов'язані з мітохондріями, органели, пов'язані з подвійною мембраною, які виробляють молекулярний водень, що використовується в клітинному метаболізмі. Вчені вважають, що гідрогеносоми, як і мітосоми, теж еволюціонували з мітохондрій. 5
Плазмові мембрани
Плазмова мембрана еукаріотичних клітин схожа за структурою з прокаріотичної плазматичної мембраною тим, що складається переважно з фосфоліпідів, що утворюють бішар з вбудованими периферичними і інтегральними білками (рис.\(\PageIndex{17}\)). Ці мембранні компоненти рухаються в площині мембрани відповідно до моделі мозаїки рідини. Однак, на відміну від прокаріотичної мембрани, еукаріотичні мембрани містять стерини, включаючи холестерин, які змінюють плинність мембрани. Крім того, багато еукаріотичних клітин містять деякі спеціалізовані ліпіди, включаючи сфінголіпіди, які, як вважають, відіграють певну роль у підтримці стабільності мембрани, а також беруть участь у шляхах трансдукції сигналів та зв'язку між клітинами.

Мембранні транспортні механізми
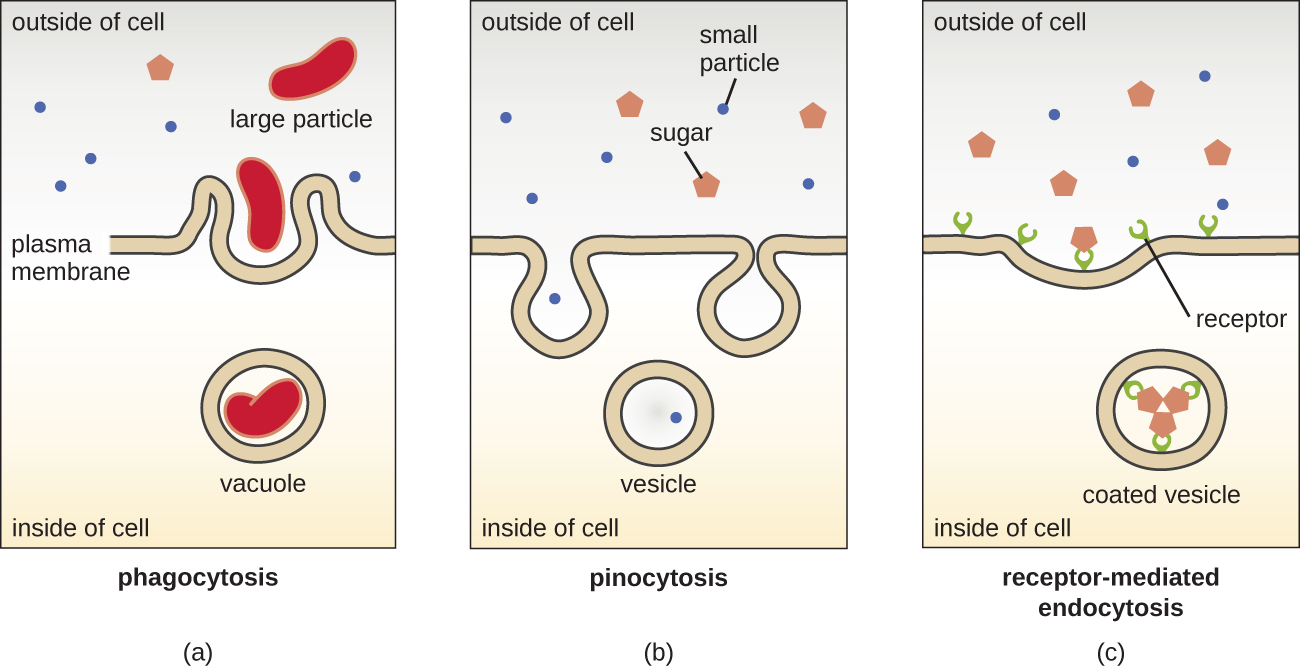
Процеси простої дифузії, полегшеної дифузії та активного транспорту використовуються як в еукаріотичних, так і в прокаріотичних клітині. Однак еукаріотичні клітини також мають унікальну здатність виконувати різні види ендоцитозу, поглинання речовини через інвагінацію плазматичної мембрани і формування вакуолі/везикул (рис.\(\PageIndex{18}\)). Тип ендоцитозу, що включає поглинання великих частинок через мембранну інвагінацію, називається фагоцитозом, що означає «поїдання клітин». При фагоцитозі частинки (або інші клітини) укладені в кишеню всередині мембрани, яка потім відщипується від мембрани, утворюючи вакуоль, яка повністю оточує частинку. Ще один вид ендоцитозу називається піноцитоз, що означає «пиття клітин». При піноцитозі дрібні розчинені матеріали і рідини забираються в клітку через дрібні бульбашки. Наприклад, сапрофітні гриби отримують свої поживні речовини з мертвих і гниючих речовин значною мірою через піноцитоз.
Ендоцитоз, опосередкований рецепторами, - це тип ендоцитозу, який ініціюється специфічними молекулами, які називаються лігандами, коли вони зв'язуються з рецепторами поверхні клітин на мембрані. Ендоцитоз, опосередкований рецепторами, - це механізм, який пептидні та амінні гормони використовують для потрапляння в клітини, а також використовується різними вірусами та бактеріями для потрапляння в клітини-господаря.
Процес, за допомогою якого секреторні бульбашки виділяють свій вміст до зовнішньої сторони клітини, називається екзоцитозом. Везикули рухаються до плазматичної мембрани і потім зливаються з мембраною, викидаючи їх вміст з клітини. Екзоцитоз використовується клітинами для видалення продуктів життєдіяльності, а також може використовуватися для випуску хімічних сигналів, які можуть бути прийняті іншими клітинами.
Клітинна стінка
Крім плазматичної мембрани, деякі еукаріотичні клітини мають клітинну стінку. Клітини грибів, водоростей, рослин і навіть деяких протестів мають клітинні стінки. Залежно від типу еукаріотичної клітини, клітинні стінки можуть бути виготовлені з широкого спектру матеріалів, включаючи целюлозу (гриби та рослини); біогенний кремнезем, карбонат кальцію, агар та карагенан (протисти та водорості); або хітин (гриби). Загалом, всі клітинні стінки забезпечують структурну стабільність клітини та захист від стресів навколишнього середовища, таких як висихання, зміни осмотичного тиску та травматичні ушкодження. 6
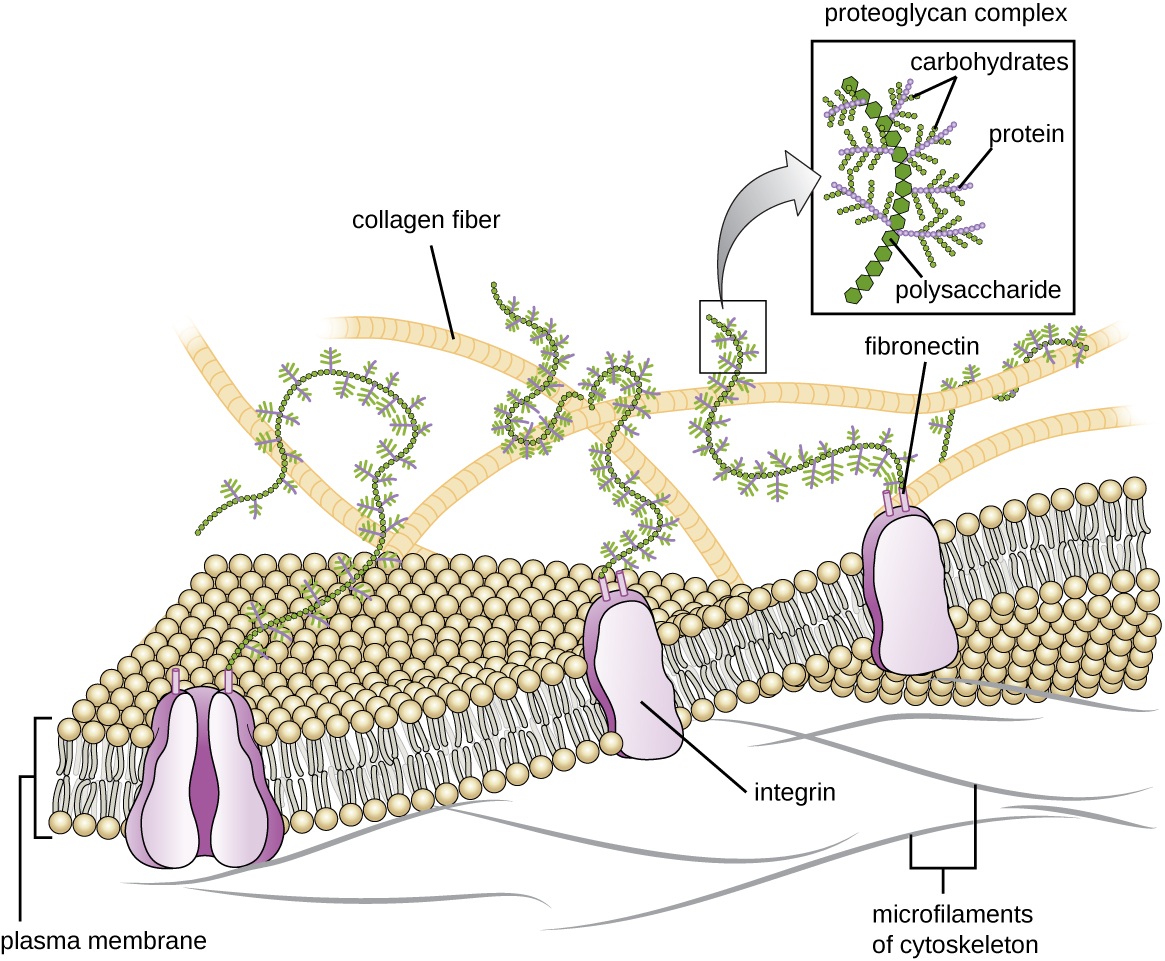
Позаклітинний матрикс
Клітини тварин і деяких найпростіших не мають клітинних стінок, щоб допомогти зберегти форму і забезпечити структурну стійкість. Натомість ці типи еукаріотичних клітин виробляють для цієї мети позаклітинний матрикс. Вони виділяють липку масу вуглеводів і білків в проміжки між сусідніми клітинами (рис.\(\PageIndex{19}\)). Деякі білкові компоненти збираються в базальну мембрану, до якої прилипають інші компоненти позаклітинного матриксу. Протеоглікани зазвичай утворюють громіздку масу позаклітинного матриксу, тоді як волокнисті білки, як колаген, забезпечують міцність. І протеоглікани, і колаген приєднуються до білків фібронектину, які, в свою чергу, прикріплюються до білків інтегрину. Ці білки інтегрину взаємодіють з трансмембранними білками в плазматичних мембранах еукаріотичних клітин, яким не вистачає клітинних стінок.
У клітині тварин позаклітинний матрикс дозволяє клітинам всередині тканин протистояти зовнішнім навантаженням і передає сигнали з зовнішньої сторони клітини всередину. Кількість позаклітинного матриксу досить велике в різних типах сполучних тканин, а варіації позаклітинного матриксу можуть надавати різним типам тканин свої відмінні властивості. Крім того, позаклітинний матрикс клітини-господаря часто є місцем, де мікробні патогени приєднуються для встановлення інфекції. Наприклад, Streptococcus pyogenes, бактерія, яка викликає стрептокок в горлі та різні інші інфекції, зв'язується з фібронектином у позаклітинному матриксі клітин, що вистилають ротоглотку (верхня область горла).
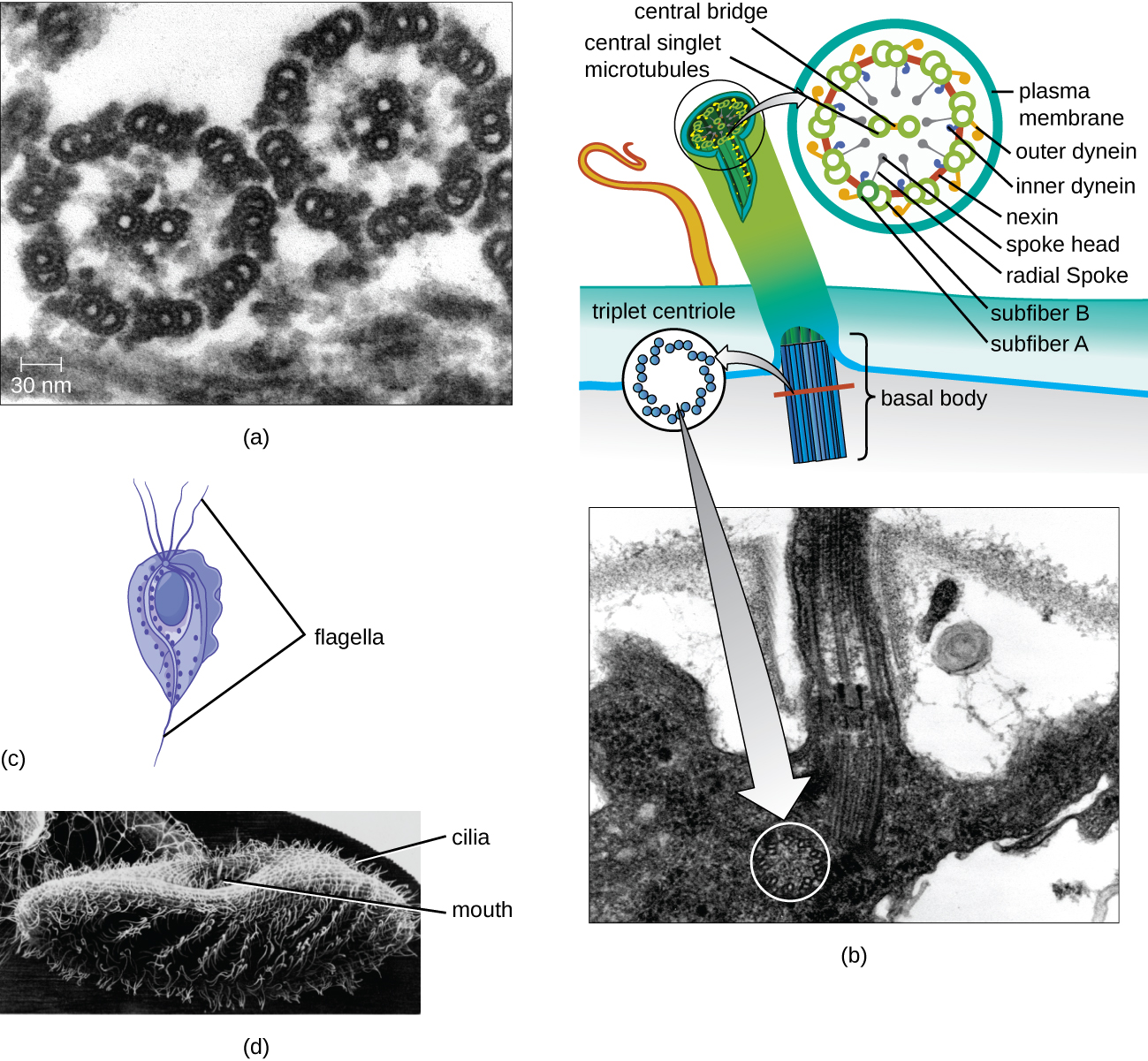
Джгутики і вії
Деякі еукаріотичні клітини використовують джгутики для локомоції; однак еукаріотичні джгутики структурно відрізняються від тих, що містяться в прокаріотичних клітині. Тоді як прокаріотичний джгутик являє собою жорстку, що обертається структуру, еукаріотичний джгутик більше схожий на гнучкий батіг, що складається з дев'яти паралельних пар мікротрубочок, що оточують центральну пару мікротрубочок. Таке розташування іменується масивом 9+2 (рис.\(\PageIndex{20}\)). Паралельні мікротрубочки використовують рухові білки диненина для переміщення відносно один одного, змушуючи джгутик згинатися.
Вії (сингулярні: війки) - це подібна зовнішня структура, виявлена в деяких еукаріотичних клітині. Унікальні для еукаріотів вії коротше джгутиків і часто покривають всю поверхню клітини; однак вони структурно схожі на джгутики (масив мікротрубочок 9+2) і використовують той же механізм для руху. Будова, яка називається базальним тілом, знаходиться біля основи кожної вії і джгутика. Базальне тіло, яке прикріплює вію або джгутик до клітини, складається з масиву триплетних мікротрубочок, подібних до центріоли, але вбудованих в плазмову мембрану. Через їх меншої довжини вії використовують швидке, гнучке, що махає рух. Крім рухливості, вії можуть мати інші функції, такі як підмітання частинок повз або в клітини. Наприклад, війчасті найпростіші використовують підмітання вій для переміщення частинок їжі в ротовий апарат, а війчасті клітини дихальних шляхів ссавців синхронно б'ються, щоб змітати слиз і сміття вгору і з легенів (рис.\(\PageIndex{20}\)).
Вправа\(\PageIndex{4}\)
- Поясніть, як клітинна оболонка еукаріотичних клітин порівнюється з оболонкою прокаріотичних клітин.
- Поясніть різницю між еукаріотичними і прокаріотичними джгутиками.
Клінічна спрямованість: Дозвіл
Оскільки амоксицилін не вирішив випадок пневмонії Барбари, ПА призначає інший антибіотик - азитроміцин, який націлений на бактеріальні рибосоми, а не на пептидоглікан. Після прийому азитроміцину за вказівкою симптоми Барбари усуваються, і вона нарешті знову починає відчувати себе. Припускаючи, що лікарська стійкість до амоксициліну не була задіяна, а з огляду на ефективність азитроміцину, збудником барбарської пневмонії, швидше за все, є Mycoplasma pneumoniae. Незважаючи на те, що ця бактерія є прокаріотичною клітиною, вона не інгібується амоксициліном, оскільки вона не має клітинної стінки і, отже, не робить пептидоглікан.
Ключові поняття та резюме
- Еукаріотичні клітини визначаються наявністю ядра, що містить геном ДНК і пов'язаного ядерною мембраною (або ядерною оболонкою), що складається з двох ліпідних бішарів, які регулюють транспорт матеріалів всередину і з ядра через ядерні пори.
- Морфологія клітин еукаріот сильно варіюється і може підтримуватися різними структурами, включаючи цитоскелет, клітинну мембрану та/або клітинну стінку.
- Ядро, розташоване в ядрі еукаріотичних клітин, є місцем рибосомального синтезу і перших етапів збірки рибосом.
- Еукаріотичні клітини містять рибосоми 80S в грубому ендоплазматичному ретикулумі (мембранні обмежені-рибосоми) і цитоплазмі (вільні рибосоми). Вони містять рибосоми 70-х років в мітохондріях і хлоропластах.
- Еукаріотичні клітини розвинули ендомембранну систему, що містить мембранозв'язані органели, що беруть участь у транспорті. До них відносяться везикули, ендоплазматичний ретикулум і апарат Гольджі.
- Гладка ендоплазматична сітка відіграє певну роль у біосинтезі ліпідів, вуглеводному обміні та детоксикації токсичних сполук. Грубий ендоплазматичний ретикулум містить мембранозв'язані рибосоми 80S, які синтезують білки, призначені для клітинної мембрани.
- Апарат Гольджі обробляє білки та ліпіди, як правило, за рахунок додавання молекул цукру, виробляючи глікопротеїни або гліколіпіди, компоненти плазматичної мембрани, які використовуються для зв'язку між клітиною.
- Лізосоми містять травні ферменти, які розщеплюють дрібні частинки, що потрапляють в організм при ендоцитозі, великі частинки або клітини, що потрапляють в організм при фагоцитозі, і пошкоджені внутрішньоклітинні компоненти.
- Цитоскелет, що складається з мікрофіламентів, проміжних ниток і мікротрубочок, забезпечує структурну підтримку в клітинах-еукаріотів і служить мережею для транспортування внутрішньоклітинних матеріалів.
- Центросоми є мікротрубочкоорганізуючими центрами, важливими для формування мітотичного веретена при мітозі.
- Мітохондрії є місцем клітинного дихання. Вони мають дві мембрани: зовнішню мембрану і внутрішню мембрану з кристами. Мітохондріальний матрикс всередині внутрішньої мембрани містить мітохондріальну ДНК, рибосоми 70S та метаболічні ферменти.
- Плазмова мембрана еукаріотичних клітин структурно схожа на ту, що міститься в прокаріотичних клітині, а мембранні компоненти рухаються відповідно до моделі рідкої мозаїки. Однак еукаріотичні мембрани містять стерини, які змінюють плинність мембран, а також глікопротеїни та гліколіпіди, які допомагають клітині розпізнавати інші клітини та інфекційні частинки.
- Окрім активного транспорту та пасивного транспорту, мембрани клітин еукаріотів можуть приймати матеріал у клітину за допомогою ендоцитозу або виганяти речовину з клітини за допомогою екзоцитозу.
- Клітини грибів, водоростей, рослин і деяких протестів мають клітинну стінку, тоді як клітини тварин і деяких найпростіших мають липкий позаклітинний матрикс, який забезпечує структурну підтримку і опосередковує клітинну сигналізацію.
- Еукаріотичні джгутики структурно відрізняються від прокаріотичних джгутиків, але служать аналогічній меті (локомоції). Вії структурно схожі на еукаріотичні джгутики, але коротші; їх можна використовувати для пересування, харчування або переміщення позаклітинних частинок.
Виноски
- 1 А.Е. Барнхілл, М.Т. Брюер, С.А. Карлсон. «Побічні ефекти антимікробних препаратів за допомогою передбачуваного або своєрідного інгібування мітохондріальних компонентів господаря». Антимікробні засоби та хіміотерапія 56 № 8 (2012) :4046—4051.
- 2 Фукс Е, Клівленд DW. «Структурні риштування проміжних ниток у здоров'ї та хворобах». Наука 279 № 5350 (1998): 514—519.
- 3 Е. Фукс, Д.В. Клівленд. «Структурні риштування проміжних ниток у здоров'ї та хворобах». Наука 279 № 5350 (1998): 514—519.
- 4 Е. Фукс, Д.В. Клівленд. «Структурні риштування проміжних ниток у здоров'ї та хворобах». Наука 279 № 5350 (1998): 514—519.
- 5 Ярлетт, Дж.Х.П. Хакштейн. «Гідрогеносоми: одна органела, множинне походження». Біонаука 55 № 8 (2005): 657—658.
- Дудзик М. «Протисти». OpenStax CNX. 27 листопада 2013 року. http://cnx.org/contents/f7048bb6-e46...ef291cf7049c@1