1.3: Коротко, генетика




- Last updated
- Oct 24, 2022
- Save as PDF

Деякі основні терміни
ДНК - це дволанцюговий полімер нуклеїнової кислоти, який є первинним спадковим матеріалом у більшості живих організмів. Кожна нитка ДНК складається з ковалентно зв'язаних нуклеотидів. Кожен нуклеотид складається з цукру, фосфату та азотистої основи. Ковалентний зв'язок між нуклеотидами відбувається між цукром одного нуклеотиду і фосфатом наступного. У ДНК є один тип цукру, але 4 типи основ. Ці основи - аденін, тимін, цитозин і гуанін - дають кожному нуклеотиду свою назву (скорочено A, T, C і G).
Дві нитки ДНК пов'язані між собою водневими зв'язками між азотистими підставами. Кожна азотиста основа має «комплементарну» основу. Пари з T, і C пари з G. Якщо ми думаємо про нитку ДНК, що має орієнтацію цукор-фосфат-цукор-фосфат-цукор-фосфат-цукор-фосфат, то друга нитка молекули має орієнтацію фосфат-цукор-фосфат-цукор-фосфат-цукор. Ми називаємо цей тип орієнтації антипаралельним, і ми називаємо ці дві пасма зворотними доповненнями. Зворотна комплементарність відноситься як до їх протилежної орієнтації (зворотної), так і до шаблону склеювання A=T G=C (доповнення).
РНК має подібну структуру, хоча зазвичай зустрічається як одна нитка. Він здатний утворювати подвійні нитки (або навіть більш складні структури, наприклад тРНК) за допомогою водневого зв'язку зі зворотними послідовностями комплементу. Він може зв'язуватися з собою або з іншими молекулами РНК.
Центральна догма молекулярної біології
Здебільшого, коли ми думаємо про молекулярну генетику, ми думаємо про створення функціональних білків з гена, закодованого в ДНК, хоча деяка РНК функціонує сама по собі.
Виробництво білка з коду ДНК відбувається за допомогою двох високо регульованих процесів: транскрипція використовує РНК-полімеразу, щоб зробити одну багатониткову молекулу мРНК з одного нитка подвійної багатониткової молекули ДНК. Переклад використовує рибосому, щоб зробити пептид (частина або весь білок) з мРНК (рис. 1).
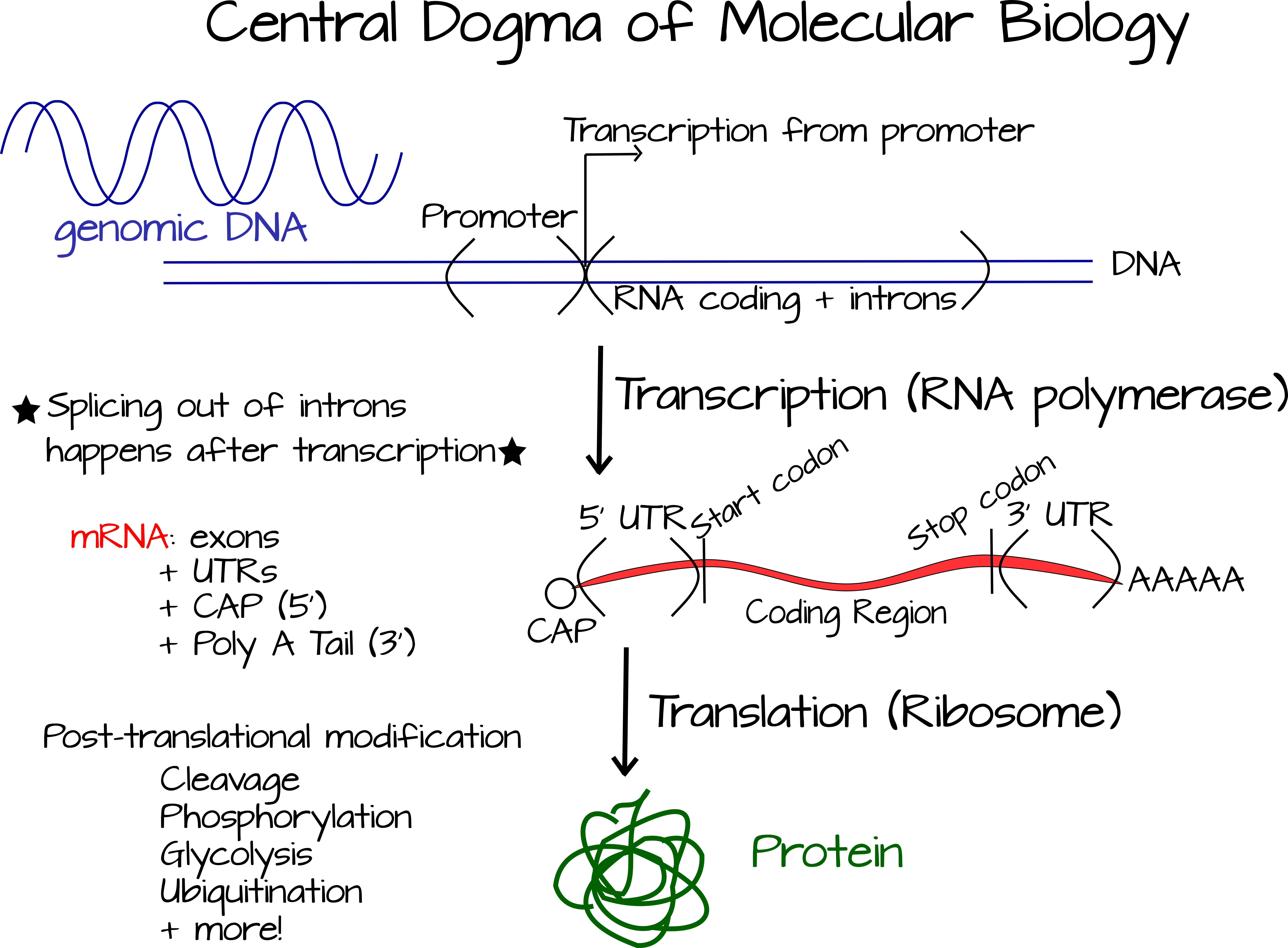
Швидка сторона примітка, тому що багато людей плутають деякі терміни, які ми використовуємо в генетиці, еволюції та розвитку. Ген відноситься до шматочка ДНК, який кодує РНК. Ця РНК часто є мРНК, але також може бути тРНК, рРНК, SNRNA, miRNA або іншим «некодуючим» типом РНК. мРНК - це РНК, яка кодує білок. Більшу частину часу «експресія генів» стосується виробництва мРНК та/або білків, для яких вони кодують. Експресія генів - це процес використання ДНК для створення функціонального продукту. Як ви, напевно, знаєте, більшість клітин у вашому організмі генетично ідентичні (більш-менш); вони «поділяють геном». Однак ваші клітини мають дуже різні функції. Ці різні функції керуються різними середовищами та різними профілями експресії генів - тобто клітини експресують різні гени. Наприклад, ми можемо очікувати знайти багато білка міозину в м'язових клітині, але багато білка Aquaporin в клітині нирок.
Регуляція експресії генів (епігенетика)
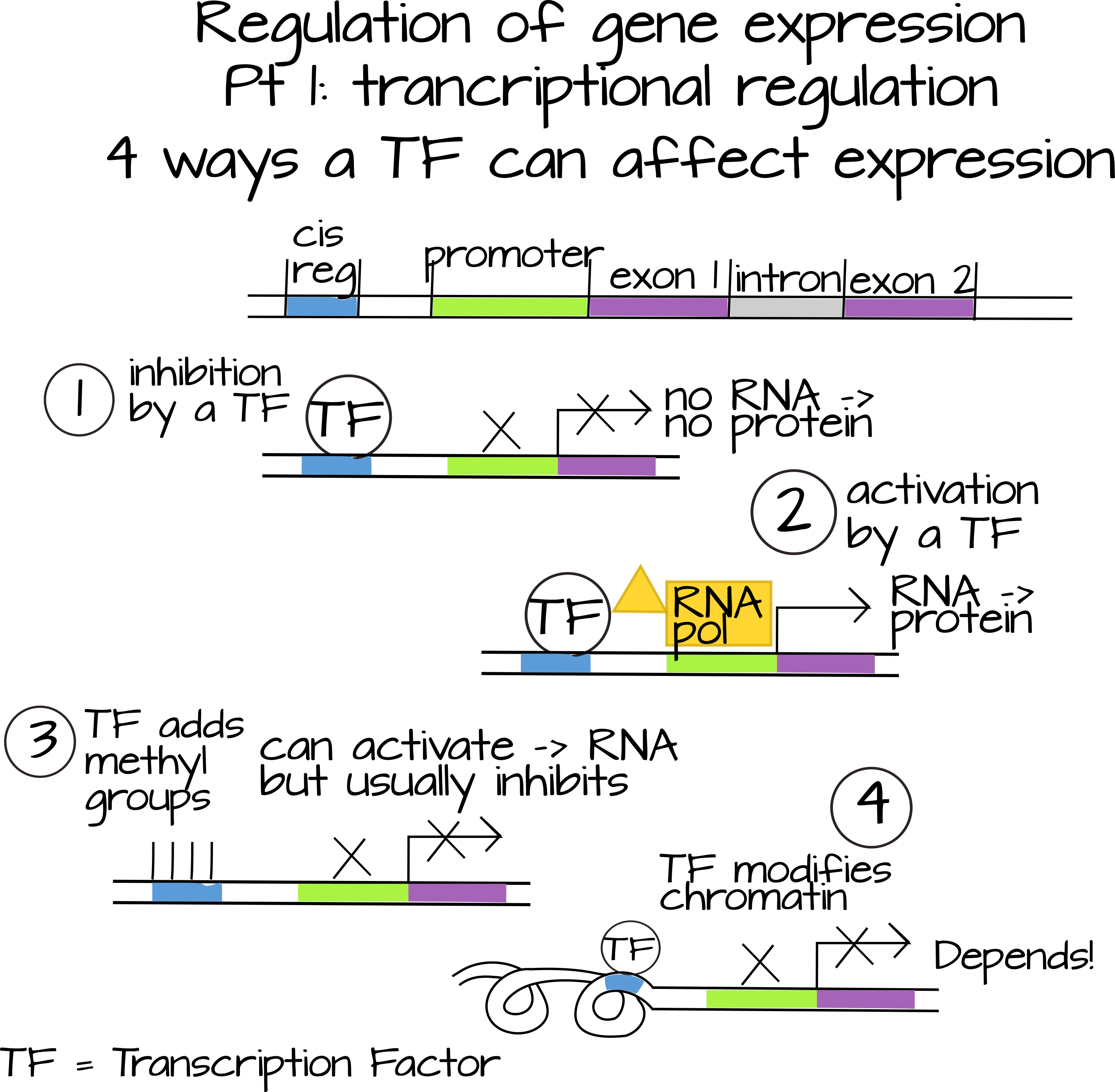
Експресія генів регулюється на трьох основних рівнях (які можна розділити на багато підрівнів). Сама транскрипція може регулюватися (ми часто називаємо це включенням гена «увімкнено» або «вимкнено»). Регулювання транскрипції - це робота факторів транскрипції, також відомих як «транс-діючі» фактори. Це дифузні молекули, часто білки, які діють різними способами, щоб збільшити або зменшити швидкість транскрипції конкретного гена. Деякі фактори транскрипції зв'язуються безпосередньо з цис-регуляторними послідовностями, шматочками ДНК, як правило, поблизу кодуючої області гена, і впливають на здатність РНК-полімерази зв'язуватися з промотором гена. Вони можуть зробити це шляхом безпосередньої взаємодії з білками, які беруть участь в наборі РНК-полімерази (наприклад, TFIID і Mediator), або шляхом зміни доступності промотора (наприклад, фізичного блокування промотора, додавання або видалення метильних груп або взаємодії з гістонами і зв'язуванням гістонів білки). Деякі фактори транскрипції зв'язуються з іншими факторами транскрипції і впливають на їх здатність зв'язувати ДНК.
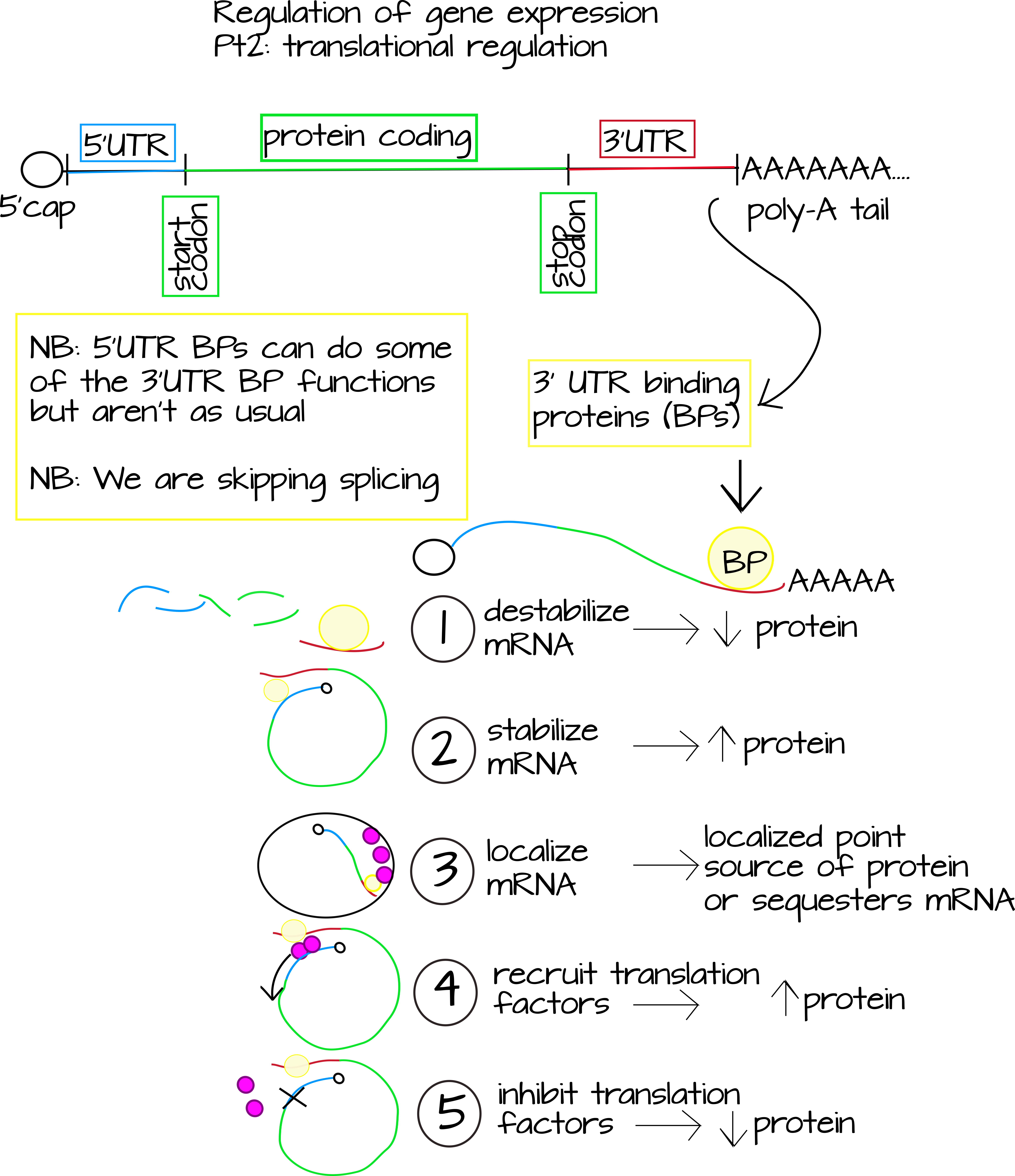
Другим основним рівнем регуляції експресії генів є поступальна регуляція. Сюди входять будь-які регулятори, які впливають на кількість білка/пептиду, що виробляється однією молекулою мРНК. Є два дуже добре вивчених способу, що це відбувається. Перший - через інтерференцію РНК (РНК), про яку ви, можливо, чули. РНК використовує короткі фрагменти некодуючої РНК, які є точними або дуже близькими зворотними доповненнями до частини послідовності мРНК. Це націлено на мРНК для руйнування або блокує трансляцію. 1 Другий добре вивчений спосіб регулювання трансляції - через РНК зв'язування білків. Ці білки часто зв'язуються зі специфічними послідовностями, виявленими поза зоною кодування білка в мРНК. Ці регіони називаються неперекладеними регіонами або «УТР». Білки, які тут зв'язуються, можуть
- Дестабілізують мРНК, знижуючи рівень трансляції.
- Стабілізувати мРНК, підвищуючи рівень трансляції.
- Локалізувати РНК до певної частини клітини.
- Блокують або гальмують зв'язування факторів перекладу, знижуючи рівні перекладу.
- Набирайте фактори перекладу, підвищуючи рівень перекладу. 2
Третім і останнім основним рівнем регулюючої експресії генів є посттрансляційний рівень. Цей рівень охоплює більшу частину того, що ми думаємо як шляхи трансдукції сигналів, або взаємодії білка, білка та малих молекул, які впливають на стабільність та функцію білка. Це може включати як ковалентну модифікацію білка (наприклад, фосфорилювання, дефосфорилювання, глікозилювання або протеолітичне розщеплення), так і нековалентну модифікацію (наприклад, інгібування або активацію аллостериків, утворення гомо і гетеродимерів). 3 Якщо ви не впевнені, що означають ці терміни зараз, це нормально - більшість з них ми не будемо охоплювати, а ті, які ми охоплюємо, легше зрозуміти в контексті, коли ви дізнаєтеся про шляхи передачі сигналу.
Посилання
- «Механізми глушіння генів дволанцюговою РНК», 2004, Природа, Гюнтер Майстер і Томас Тушль, дої:10.1038/nature02873
- «Дивний новий світ РНК-зв'язуючих білків», 2018, Відгуки природи: Молекулярна клітинна біологія, Маттіас Хенце, Альфредо Кастелло, Томас Шварцль і Томас Прайс дої:10.1038/nrm.2017.130
- «Три звички високоефективних сигнальних шляхів: принципи транскрипційного контролю за допомогою сигналізації клітин розвитку», 2002, Гени та розвиток, Скотт Бароло та Джеймс Посаконі, 10.1101/gad.976502