7.7: Мейоз
- Page ID
- 5221

Мітоз виробляє дві клітини з такою ж кількістю хромосом, що і батьківська клітина. Мітоз диплоїдної клітини (2n) продукує дві диплоїдні дочірні клітини. Якщо дві диплоїдні клітини продовжували брати участь у статевому розмноженні, їх злиття призвело б до тетраплоїдної (4n) зиготи. Рішенням цієї проблеми є Мейоз.
Мейоз
Мейоз - це процес поділу клітин у еукаріот, що характеризується:
- два послідовних поділу: мейоз I і мейоз II
- відсутність синтезу ДНК (без S-фази) між двома відділами
- результат: 4 клітини з половиною числа хромосом стартової клітини, наприклад, 2n → n
Злиття двох таких клітин виробляє 2n зиготу.
Мейоз у тварин
Використовується для вироблення гамет: сперми та яйцеклітини
Мейоз у рослин
Використовується для отримання спор. Спори - це початок генерації гаметофітів, які з часом вироблять гамети (мітозом, оскільки стартові клітини вже гаплоїдні).
Мейоз I
Профаза мейозу I (профаза I) є більш складним процесом, ніж профаза мітозу (і зазвичай займає набагато більше часу).
Ось короткий огляд процесу. Більш детальний вигляд наведено нижче.
- Коли хромосоми вперше стають видимими, вони вже подвоюються, кожен гомолог дублюється під час попередньої S-фази.
- Результат: пари гомологічних діад кожна діада, що складається з двох сестринських хроматидів, утримуваних разом білковим комплексом, званим когезином.
- Спарювання: Кожна пара гомологічних діад вирівнюється по довжині один з одним.
- Результат: тетрада. (Ці структури іноді називають двовалентними, оскільки на цьому етапі ви не можете розрізнити окремі сестринські хроматиди під мікроскопом.)
- Дві гомологічні діади утримуються
- одна або кілька хіазмат (співати. = хіазма), які утворюються між двома несестринними хроматидами в точках, де вони перетиналися.
- синаптонемальний комплекс (СК), складна збірка білків (включаючи когезин)
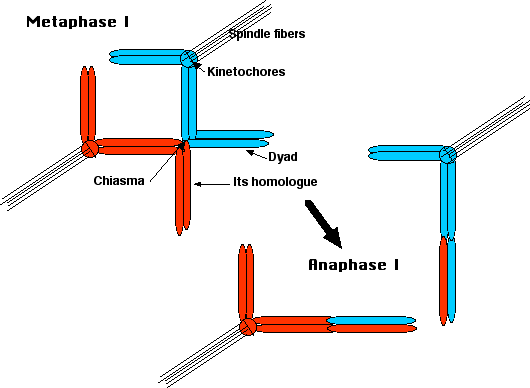
При метафазі I мікротрубочки волокон веретена прикріплюються до
- сестринські кінетохори одного гомолога, що тягнуть обидві сестринські хроматиди до одного полюса клітини;
- Сестра кінетохорів іншого гомологу тягне цих сестер до протилежного полюса.
Результат: один гомолог витягується вище метафазної пластини, інший внизу. Хіазмати тримають гомологів, прикріплених один до одного, а когезин тримає сестринських хроматидів разом.
При анафазі я,
- когезин між плечима хромосоми руйнується, дозволяючи
- хіазмати, щоб розсунути.
- Результат: гомологічні діади відокремлюються і мігрують до відповідних полюсів.
Мейоз II
Поведінка хромосом при мейозі II подібна до мітозу.
- При метафазі II шпиндельні волокна прикріплюють один кінетохора діади до одного полюса, інший - до протилежного полюса.
- При анафазі II хроматиди відокремлюються і (кожен тепер є незалежною хромосомою) рухаються до відповідних полюсів.
Генетична рекомбінація
Мейоз не тільки зберігає розмір генома статево відтворюваних еукаріотів, але також забезпечує три механізми для диверсифікації геномів потомства.
Перетин над
Chiasmata представляють точки, де раніше (і невидимі) несестринські хроматиди обмінялися місцями розділів. Процес називається перехрещуванням. Це взаємно; сегменти, які обмінюються кожним несестровим хроматидом, ідентичні (але можуть нести різні алелі).
Кожен хроматид містить єдину молекулу ДНК. Отже, проблема перетину насправді є проблемою обміну частинами сусідніх молекул ДНК. Це потрібно робити з великою точністю, щоб ні хроматид не здобув і не втрачав жодних генів. Насправді перетин повинен бути достатньо точним, щоб жоден нуклеотид не втрачався або не додавався в точці кросовера, якщо це відбувається всередині гена. В іншому випадку це призведе до зрушення кадру, і отриманий ген буде виробляти дефектний продукт або, швидше за все, взагалі жоден продукт.

На схемі вище показана тільки одинична хіазма. Однак зазвичай зустрічаються множинні хіазмати (у людини середня кількість хіазмат на тетрада становить трохи більше двох). На цій фотографії тетрада коника Chorthippus parallelus показує 5 хіазмат.
Випадковий асортимент
При мейозі I орієнтація батьківських і материнських гомологів у метафазної пластини випадкова. Тому, хоча кожна клітина, вироблена мейозом, містить тільки один з кожного гомологу, кількість можливих поєднань материнських і батьківських гомологів становить 2 n, де n = гаплоїдне число хромосом. На цій схемі гаплоїдне число дорівнює 3, а 8 (2 3) випускаються різні комбінації.
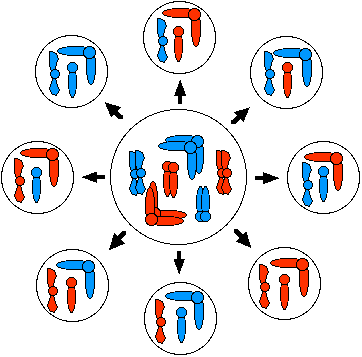
Випадковий асортимент гомологів у людини виробляє 2 23 (8 388 608) різних комбінацій хромосом.
Крім того, через схрещування жодна з цих хромосом не є «чистою» материнською чи батьківською. Розподіл рекомбінантних і нерекомбінантних сестринських хроматидів в дочірні клітини при анафазі II також є випадковим.
Тому я думаю, що можна з упевненістю зробити висновок, що з усіх мільярдів сперматозоїдів, вироблених чоловіком за його життя (і сотні яйцеклітин, які дозрівають протягом життя жінки), жоден з двох не має абсолютно однакового вмісту генів.
Запліднення
Зменшуючи кількість хромосом з 2n до n, встановлюється стадія об'єднання двох геномів. Якщо батьки розрізняються генетично, у їх потомства можуть виникати нові комбінації генів.
Беручи ці три механізми разом, я думаю, що можна з упевненістю зробити висновок, що жодна людина ніколи не поділяла однакового генома, якщо вони не мали однакового брата; це брат, що виробляється з тієї ж заплідненої яйцеклітини.
Поведінка хромосом при мейозі (2n → n) і заплідненні (n + n → 2n) забезпечують структурну основу правил спадкування Менделя.
Prophase I — детальний перегляд
Тривалі та складні події профази I можна розбити на 5 етапів.
Лептотен
- Всі хромосоми ущільнюються.
- Спарювання. Гомологічні діади (пари сестринських хроматидів) знаходять один одного і вирівнюються від кінця до кінця за допомогою осьового елемента (який містить когезин). У бутонізації дріжджів (і, можливо, інших еукаріотів) процес слідує періоду випробувань і помилок. Будь-які дві діади пари в своїх центромерах. Якщо вони не гомологи, вони відокремлюються і намагаються ще раз.
- Як несестри розпізнають свої спільні області гомології ДНК, є невизначеним. Дволанцюгові розриви (DSBs) часто трапляються в ДНК хроматидів, і вони можуть бути необхідні для того, щоб гомологи розпізнали один одного.
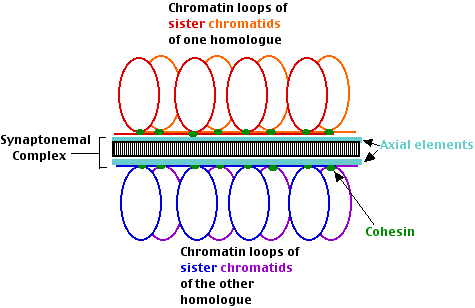
Зиготен
- Синапсис. Починає формуватися синаптонемальний комплекс.
- ДНК-нитки несестринних хроматидів починають процес рекомбінації. Як вони здатні це зробити через синаптонемальний комплекс, який має товщину понад 100 нм, невідомо.
Пахітене
- Синапсис зараз завершений.
- З'являються рекомбінаційні вузлики (по крайней мере, у деяких організмів, в тому числі і у людини). Вони названі за ідею, що вони представляють точки, де відбувається рекомбінація ДНК.
- Для кожного двовалентного має бути принаймні один, якщо мейоз має досягти успіху. Їх часто більше, кожен з них імовірно представляє точку кросовера.
- Вони містять ферменти, які, як відомо, необхідні для рекомбінації та відновлення ДНК.
- Етапи рекомбінування ДНК тривають до кінця пахітену.
Диплотен
- Рекомбінація ДНК завершена.
- Починає руйнуватися синаптонемальний комплекс.
- Хроматиди починають відриватися, виявляючи
- хіазмати. Спочатку хіазмати розташовуються в місцях рекомбінаційних вузликів, але пізніше вони мігрують до кінців хроматид.
Діакінез
У деяких організмів хромосоми деконденсируются і починають транскрибуватися на деякий час. Далі відбувається реконденсація хромосоми при підготовці до метафази I.
У істот, де цього не відбувається, хромосоми конденсуються далі при підготовці до метафази I.
Контроль якості мейозу
Не повинно дивуватися, що в такому складному процесі все може піти не так. Однак клітини, що проходять через мейоз, мають контрольно-пропускні пункти, які контролюють кожну пару гомологів для
- правильна рекомбінація їх ДНК
- правильне формування синаптонемального комплексу
Будь-який виявлений збій зупиняє процес і зазвичай змушує клітину самознищуватися апоптозом.
Однак, незважаючи на ці контрольні точки, помилки іноді залишаються невиправленими.
Помилки при мейозі
Підраховано, що з 10— 25% всіх запліднених яйцеклітин людини містять хромосомні аномалії, і це найпоширеніша причина недостатності вагітності (35% випадків).
Ці хромосомні аномалії
- виникають через помилки при мейозі, зазвичай мейозі I;
- виникають частіше (90%) під час утворення яйцеклітини, ніж при утворенні сперматозоїдів;
- частішають у міру старіння жінки.
- Аневплоїдія - посилення або втрата цілих хромосом - є найпоширенішою хромосомною аномалією. Вона викликана нераз'єктированностью, нездатністю хромосом правильно відокремлюватися:
- гомологи під час мейозу I або
- сестринські хроматиди під час мейозу II
- Зиготи, що не мають однієї хромосоми («моносомія»), не можуть розвинутися до народження (за винятком самок з єдиною Х-хромосомою).
- Три однакові хромосоми («трисомія») також смертельні за винятком хромосом 13, 18 і 21 (трисомія 21 є причиною синдрому Дауна).
- Три або більше Х-хромосоми життєздатні, оскільки всі, крім однієї, інактивовані.