7.2: Клітинний цикл
- Page ID
- 5207

Еукаріотична клітина не може розділитися на дві, дві на чотири тощо, якщо не чергуються два процеси:
- подвоєння його генома (ДНК) в S-фазі (фазі синтезу) клітинного циклу;
- зменшення вдвічі цього генома під час мітозу (M фаза).
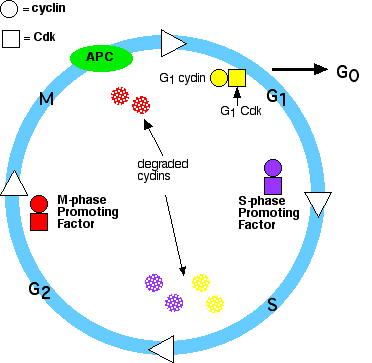
Контроль клітинного циклу
Проходження клітини через клітинний цикл контролюється білками в цитоплазмі. Серед основних гравців у тваринних клітинок:
- Їх рівні в клітині підвищуються і падають зі стадіями клітинного циклу.
- Їх рівні в клітці залишаються досить стабільними, але кожен повинен пов'язувати відповідний циклін (рівні якого коливаються), щоб активуватися. Вони додають фосфатні групи до різноманітних білкових субстратів, які контролюють процеси в клітинному циклі.
- Комплекс, що сприяє анафазі (APC). (БТР також називають циклосомою, а комплекс часто позначається як APC/C.) АТР/С
- запускає події, що призводять до руйнування когезину (як описано нижче), тим самим дозволяючи сестринським хроматидам відокремлюватися
- погіршує мітотичні (B) цикли
Етапи в циклі
- Підвищення рівня G 1 -циклінів зв'язується зі своїми Cdks і сигналізує клітині підготувати хромосоми до реплікації.
- Підвищення рівня фактора, що сприяє S-фазі (SPF) - який включає цикліни A, пов'язані з Cdk2, потрапляє в ядро і готує клітину до дублювання своєї ДНК (та її центросом).
- У міру продовження реплікації ДНК циклін Е руйнується, і рівень мітотичних циклінів починає підвищуватися (в G 2).
- Ініціює транслокацію М-фазового стимулюючого фактора (комплексу мітотичних циклінів [B] з М-фазою Cdk [Cdk1]) в ядро
- збірка мітотичного веретена
- розрив ядерної оболонки
- припинення транскрипції всіх генів
- конденсація хромосом
- Ці події призводять клітину до метафази мітозу.
- У цей момент фактор, що сприяє M-фазі, активує комплекс, що сприяє анафазі (APC/C), який
- дозволяє сестринським хроматидам у метафазної пластини відокремлюватися і рухатися до полюсів (= анафази), завершуючи мітоз.
Поділ сестринських хроматидів залежить від розпаду когезину, який утримував їх разом. Це працює так.
- Розпад когезину викликаний протеазою, яка називається сепараза (також відома як сепарин).
- Сепараза зберігається неактивною до пізньої метафази інгібуючим шапероном, який називається секурін.
- Анафаза починається, коли комплекс, що сприяє анафазі (APC/C), руйнує securin (шляхом позначення його убіквітином для депозиту в протеасомі), тим самим закінчуючи його гальмування сепарази і дозволяючи
- відокремити, щоб розбити когезин
- руйнує цикли B. Це також робиться шляхом приєднання їх до убіквітину, який націлений на них для знищення протеасомами.
- включає синтез циклів G 1 (D) на наступний виток циклу.
- деградує гемінін, білок, який утримував свіжосинтезовану ДНК у S-фазі від повторної реплікації до мітозу.
Це лише один із механізмів, за допомогою якого клітина гарантує, що кожна частина її генома копіюється один раз - і лише один раз - під час S-фази.
Деякі клітини навмисно скорочують клітинний цикл коротко, дозволяючи повторювати S-фази без завершення мітозу та/або цитокінезу. Це називається ендореплікацією.
Мейоз і клітинний цикл
Особлива поведінка хромосом при мейозі I вимагає деякого особливого контролю. Тим не менш, проходження клітинного циклу при мейозі I (а також мейозі II, який по суті є мітотичним поділом) використовує багато однакових гравців, наприклад, MPF і APC. (Насправді, MPF також називають моїм насиченням-р сприяння фактору за його роль в мейозі I і II розвиваються яйцеклітин.
Контроль якості клітинного циклу
Клітина має кілька систем переривання клітинного циклу, якщо щось піде не так.
Контрольно-пропускні пункти пошкодження ДНК. Ці чутливі пошкодження ДНК як до того, як клітина увійде в S-фазу (контрольна точка G 1), так і після S фази (контрольна точка G 2). Пошкодження ДНК до того, як клітина увійде в S-фазу, гальмує дію Cdk2, тим самим зупиняючи прогресування клітинного циклу, поки пошкодження не можна буде відновити. Якщо пошкодження настільки серйозне, що його неможливо відновити, клітина саморуйнується апоптозом. Пошкодження ДНК після S-фази (контрольна точка G 2) пригнічує дію Cdk1, тим самим запобігаючи клітині переходити від G 2 до мітозу. Перевірка успішної реплікації ДНК під час S-фази. Якщо реплікація припиняється в будь-якій точці ДНК, прогрес через клітинний цикл зупиняється, поки проблема не буде вирішена.
Контрольні точки шпинделя. Деякі з них, які були виявлені для виявлення будь-якої несправності волокон шпинделя приєднуватися до кінетохорів і заарештувати клітину в метафазі, поки всі кінетохори не будуть прикріплені правильно. Вони виявляють неправильне вирівнювання самого веретена і блокують цитокінез. Крім того, вони викликають апоптоз, якщо шкода непоправна. Всі оглянуті контрольно-пропускні пункти вимагають послуг комплексу білків. Мутації в генах, що кодують деякі з них, були пов'язані з раком; тобто вони є онкогенами. Це не повинно дивуватися, оскільки збої контрольно-пропускних пунктів дозволяють осередку продовжувати ділитися, незважаючи на пошкодження її цілісності.
Приклади контрольних точок
p53
Білок p53 відчуває пошкодження ДНК і може зупинити прогресування клітинного циклу в G 1 (блокуючи активність Cdk2). Обидві копії гена p53 повинні бути мутовані, щоб це не вдалося, тому мутації в p53 є рецесивними, а p53 кваліфікується як ген супресора пухлини.
Білок p53 також є ключовим гравцем в апоптозі, змушуючи «погані» клітини покінчити життя самогубством. Отже, якщо клітина має лише мутантні версії білка, вона може жити далі - можливо, перерости в рак. Більше половини всіх видів раку людини насправді містять мутації p53 і не мають функціонуючого білка p53.