9.2: Черв'ячний ланцюг




- Last updated
- Oct 25, 2022
- Save as PDF
Черв'ячний ланцюг (WLC) - це, мабуть, найбільш часто зустрічаються моделі полімерного ланцюга при описі механіки і термодинаміки макромолекул. Ця модель описує поведінку тонкого гнучкого стрижня і особливо корисна для опису жорстких ланцюгів зі слабкою кривизною, таких як подвійна багатониткова ДНК. Його поведінка залежить тільки від двох параметрів, які описують стрижень:κb його жорсткість на вигин іLC, довжина контуру.
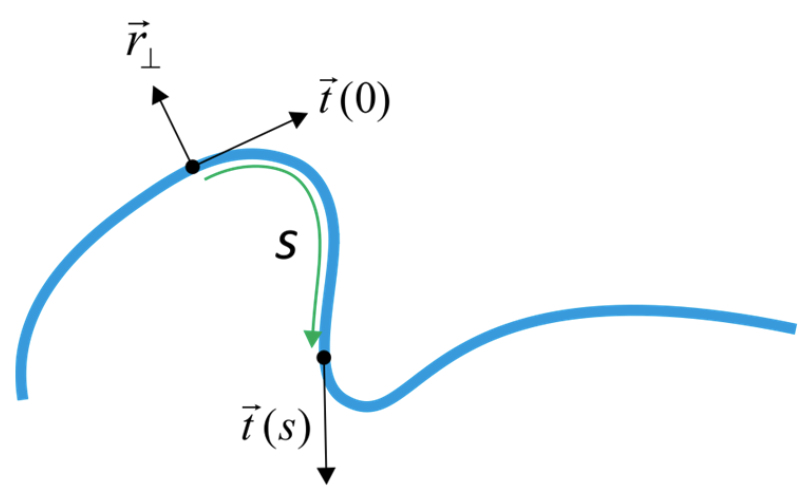
Давайте визначимо змінні в цій моделі WLC:
- sThe distance separating two points along the contour of the rod
- →r⊥Normal unit vector
- →t=∂→r⊥∂sTangent vector
- ∂→t∂s Curvature of chain
- =1Ris inverse of local radius of curvature
Черв'ячатий ланцюг характеризується:
- Довжина стійкості, яка визначається через тангенс векторної кореляційної функції:
g(s)=⟨→t(0)⋅→t(s)⟩=exp[−|s|/ℓp]
- Енергія вигину: Енергія, необхідна для згинання дотичних векторів сегмента довжини s, може бути виражена як
Ub=12κb∫L0ds(∂→t∂s)2
Енергія згинання

Оцінимо енергію вигину WLC, зробивши деякі спрощують припущення, корисні для досить жорстких стрижнів. Якщо розглядати короткі відстані, на яких кривизна невелика, тоθ≈s/R і
∂→t∂s≈dθds=1R
Тоді ми можемо висловити енергію вигину через кут:
Ub≈12sκbθ2
Зверніть увагу на схожість цього виразу з енергією, необхідною для витіснення частинки, пов'язаної в гармонійний потенціал з постійною сили k: U = ½kx 2. Енергія вигину може бути використана для отримання термодинамічних середніх. Наприклад, ми можемо обчислити дисперсію для кутів тангенса вектора як функція s (сферичні координати):
⟨θ2(s)⟩=1Qbend∫2π0dϕ∫π0dθsinθθ2e−Ub(θ)/kBT=2skBTκb
Ось ми і скористалисяsinθ≈θ. Функція перегородки при згинанні стрижня - це:
Qbend=∫2π0dϕ∫π0dθsinθe−Ub(θ)/kBT
Стійкість Довжина
Щоб описати довжину стійкості WLC, ми визнаємо, що Equation\ ref {9.2.1} можна записати якg(s)=⟨cosθ(s)⟩ і розширити це для малогоθ:
g(s)=⟨cosθ(s)⟩=⟨1−θ2(s)2+...⟩≈1−12⟨θ2(s)⟩
і з Рівняння\ ref {9.2.3} ми можемо записати:
g(s)≈1−skBTκb
Якщо порівняти це з розширенням експоненціальної в рівнянні\ ref {9.2.1}
g(s)=e−|s|/ℓp≈1−|s|ℓp
отримуємо вираз для довжини стійкості черв'ячного ланцюга
ℓp=κbkBT
Відстань від кінця до кінця
Наскрізна відстань для WLC отримується шляхом інтеграції вектора тангенса по одній довжині контуру:
→R=∫LC0ds→t(s)
Отже, дисперсія в наскрізній відстані визначається з тангенсної векторної автокореляційної функції, яку ми приймаємо, щоб мати експоненціальну форму:
⟨R2⟩=⟨R⋅R⟩=∫LC0ds∫LC0ds′⟨t(s)t(s′)⟩=∫LC0ds∫LC0ds′e−(s−s′)/ℓp
⟨R2⟩=2ℓpLC−2ℓ2p(1−e−LC/ℓp)
Розглянемо цей вираз в двох межах:
rigid:ℓp≫LC⟨R2⟩≈L2C
\text{flexible:} \qquad \ell_p \ll L_C \qquad \langle R^2 \rangle \approx 2L_C \ell_p \rightarrow n_e \ell_e^2 \rightarrow \therefore 2\ell_p = \ell_e
Яка енергія потрібна для обгортання ДНК навколо октамера гістону в нуклеосомі? Дволанцюгова ДНК - це жорсткий полімер з довжиною персистенції\ell_p \approx 50 нм, але нуклеосома має радіус ~ 4,5 нм.

Рішення
З\ell_p і k B T = 4,1 пН нм визначити жорсткість на вигин можна за допомогою:
\kappa_b = \ell_p k_BT = (50\text{ nm})(4.1 \text{ pN nm}) = 205\text{ pN nm}^2
Тоді енергія, необхідна для згинання dsDNA в один повний цикл
\begin{aligned} U_b &\cong \dfrac{\kappa_b \theta^2}{2s} \approx \dfrac{\kappa_b (2\pi)^2}{2(2\pi R)} = \dfrac{\pi \kappa_b}{R} \\ &= \dfrac{\pi(205 \text{ pn nm}^2 )}{4.5 \text{ nm}} = 143 \text{ pN nm} \\ &= 35k_BT = 15 \text{ kcal (mol loops)}^{-1} \\ &\qquad \text{or } 0.15 \text{ kcal basepair}^{-1} \end{aligned}
Механіка континууму тонкого стрижня 1
Черв'ячний ланцюг - це модель, отримана з механіки континууму тонкого стрижня. Крім вигину, тонкий стрижень схильний до інших спотворень: розтягування, скручування, написання.

Підіб'ємо підсумки енергій, необхідних для цих деформацій:
Змінні деформації:
- S: Положення уздовж контуру стрижня
- L 0: Незбурена довжина стрижня
- \vec{t}: Дотичний вектор.
- d\vec{t}/ds: викривлення
- Ω: Місцевий поворот
Енергія для спотворення стрижня дорівнює
U = U_{st} +U_b+U_{tw} \nonumber
У гармонійному наближенні для відновлювальної сили ми можемо записати ці внески як
U= \dfrac{1}{2} \int^L_{L_0} \kappa_{st} s ds \, + \dfrac{1}{2} \int^L_{L_0} \kappa_b \left( \dfrac{d\vec{t}}{ds} \right)^2 ds + \dfrac{1}{2} \int^L_{L_0} \kappa_{tw} \Omega^2 ds \nonumber
Константи сили, з репрезентативними значеннями для dsDNA, є:
Розтяжка:\kappa_{st} = \kappa_{st-entropic} + \kappa_{st-enthalpic}
\kappa_{st-entropic} \approx 3k_BT/ \ell_p L_c
Вигин: \kappa_b
\kappa_b \approx 205 \text{ pN nm}^2
Скручування:\kappa_{tw}
\kappa_{tw} \approx (86 \text{ nm})k_BT = 353 \text{ pN nm}^2
Напишіть
Додатковим спотворенням тонких стрижнів є writhe, що відноситься до зв'язаного скручування і намотування, і є важливим фактором супернамотування ДНК. Скручування стрижня може викликати в площині петлі стрижня, наприклад, як зустрічається при спробі намотати садовий шланг. Число Writhe W стрижня відноситься до кількості повних петель, зроблених стрижнем. Writhe може бути позитивним або негативним в залежності від того, чи стрижень перетинає над собою справа наліво або зліва направо. Число скручування T - це число Ω = 2π обертань стрижня, а також може бути позитивним або негативним.

Зв'язкове число L = T+W зберігається в В-формі ДНК, так що скручування може бути перетворено в writhe і навпаки. Оскільки ДНК в клітині природно негативно перекручується в нуклеосомах, топоізомерази використовуються для зміни числа зв'язування шляхом розриву та реформування хребта фосфодіефіру після розслаблення скручування. Негативно перекручена ДНК може бути перетворена в кругову ДНК шляхом локального пузиріння (розмотування в окремі нитки).
_____________________________________
- Боал Д.Х., Механіка клітини, 2-е изд. (Кембриджський університетський прес, Кембридж, Великобританія, 2012).