18.1: Архітектура імунної системи
- Page ID
- 3929

Цілі навчання
- Визначте пам'ять, первинну реакцію, вторинну відповідь та специфіку
- Розрізняють гуморальний і клітинний імунітет
- Диференціювати антигени, епітопи та гаптени
- Опишіть структуру та функцію антитіл та розмежуйте різні класи антитіл
Клінічна спрямованість: Частина 1
Олівію, однорічну дитину, привозять до відділення невідкладної допомоги батьки, які повідомляють про її симптоми: надмірний плач, дратівливість, чутливість до світла, незвичайна млявість та блювота. Лікар відчуває набряклі лімфатичні вузли в горлі та пахвах Олівії. Крім того, область живота над селезінкою набрякла і ніжна.
Вправа\(\PageIndex{1}\)
- Про що говорять ці симптоми?
- Які тести можуть бути наказані, щоб спробувати діагностувати проблему?
Адаптивний імунітет визначається двома важливими характеристиками: специфічністю і пам'яттю. Специфічність стосується здатності адаптивної імунної системи орієнтуватися на конкретні патогени, а пам'ять стосується її здатності швидко реагувати на патогени, яким вона раніше піддавалася впливу. Наприклад, коли людина одужує від вітрянки, організм виробляє пам'ять про інфекцію, яка спеціально захистить її від збудника - вірусу вітряної віспи, якщо він пізніше знову піддається впливу вірусу.
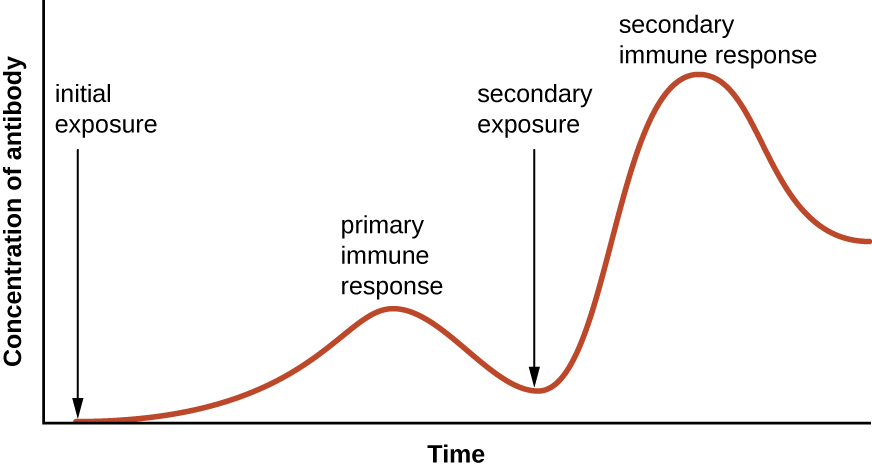
Специфічність і пам'ять досягаються, по суті, програмуванням певних клітин, що беруть участь в імунній відповіді, щоб швидко реагувати на подальше опромінення збудника. Таке програмування відбувається в результаті першого впливу збудника або вакцини, що запускає первинну відповідь. Подальші експозиції призводять до вторинної реакції, яка швидше і сильніше в результаті пам'яті організму про першу експозицію (рис.\(\PageIndex{1}\)). Однак ця вторинна відповідь специфічна для розглянутого збудника. Наприклад, вплив одного вірусу (наприклад, вірусу вітряної віспи) не забезпечить захисту від інших вірусних захворювань (наприклад, кору, свинки або поліомієліту).
Адаптивний специфічний імунітет передбачає дії двох різних типів клітин: В-лімфоцитів (В-клітин) і Т-лімфоцитів (Т-клітин). Хоча В-клітини та Т-клітини виникають із загального шляху диференціації гемопоетичних стовбурових клітин, місця їх дозрівання та їх роль у адаптивному імунітеті дуже різні.
В-клітини дозрівають в кістковому мозку і відповідають за вироблення глікопротеїнів, званих антитілами, або імуноглобулінів. Антитіла беруть участь в захисті організму від хвороботворних мікроорганізмів і токсинів у позаклітинному середовищі. Механізми адаптаційного специфічного імунітету, які залучають В-клітини і вироблення антитіл, називають гуморальним імунітетом. Дозрівання Т-клітин відбувається в тимусі. Т-клітини функціонують як центральний оркестратор як вроджених, так і адаптивних імунних реакцій. Вони також відповідають за знищення клітин, інфікованих внутрішньоклітинними збудниками. Націлювання та знищення внутрішньоклітинних збудників Т-клітинами називається клітинним імунітетом, або клітинним імунітетом.
Вправа\(\PageIndex{2}\)
- Перерахуйте дві визначальні характеристики адаптивного імунітету.
- Поясніть різницю між первинним і вторинним імунною відповіддю.
- Чим відрізняються гуморальний і клітинний імунітет?
Антигени
Активація адаптивного імунного захисту запускається патоген-специфічними молекулярними структурами, які називаються антигенами. Антигени подібні до асоційованих з патогенами молекулярних моделей (ПАМП), обговорюваних у розпізнаванні патогенів та фагоцитозі; однак, тоді як ПАМП - це молекулярні структури, виявлені на численних патогенах, антигени є унікальними для конкретного збудника. Наприклад, антигени, що стимулюють адаптивний імунітет до вітрянки, є унікальними для вірусу вітряної віспи, але значно відрізняються від антигенів, пов'язаних з іншими вірусними збудниками.
Термін антиген спочатку використовувався для опису молекул, які стимулюють вироблення антитіл; насправді цей термін походить від поєднання слів анти тіло і ген ератор, а молекула, яка стимулює вироблення антитіл, вважається антигенною. Однак роль антигенів не обмежується гуморальним імунітетом і виробленням антитіл; антигени також відіграють істотну роль у стимулюванні клітинного імунітету, і з цієї причини антигени іноді більш точно називають імуногенами. Однак у цьому тексті ми зазвичай називаємо їх антигенами.
Збудники мають різноманітними структурами, які можуть містити антигени. Наприклад, антигени з клітин бактерій можуть бути пов'язані з їх капсулами, клітинними стінками, бахромою, джгутиками або пілі. Бактеріальні антигени також можуть бути пов'язані з позаклітинними токсинами і ферментами, які вони виділяють. Віруси мають різноманітні антигени, пов'язані з їх капсидами, оболонками та спайковими структурами, які вони використовують для прикріплення до клітин.
Антигени можуть належати до будь-якої кількості молекулярних класів, включаючи вуглеводи, ліпіди, нуклеїнові кислоти, білки та комбінації цих молекул. Антигени різних класів відрізняються своєю здатністю стимулювати адаптивний імунний захист, а також за типом реакції, яку вони стимулюють (гуморальну або клітинну). Структурна складність антигенної молекули є важливим фактором її антигенного потенціалу. Взагалі більш складні молекули більш ефективні в якості антигенів. Наприклад, тривимірна складна структура білків робить їх найбільш ефективними і сильнодіючими антигенами, здатними стимулювати як гуморальний, так і клітинний імунітет. Для порівняння, вуглеводи менш складні за структурою і тому менш ефективні як антигени; вони можуть лише стимулювати гуморальний імунний захист. Ліпіди та нуклеїнові кислоти є найменш антигенними молекулами, а в деяких випадках можуть стати антигенними лише при поєднанні з білками або вуглеводами з утворенням гліколіпідів, ліпопротеїнів або нуклеопротеїнів.
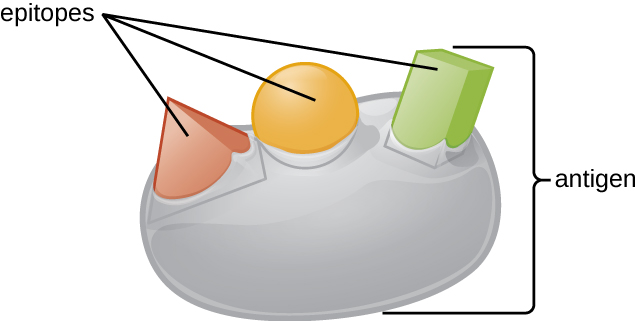
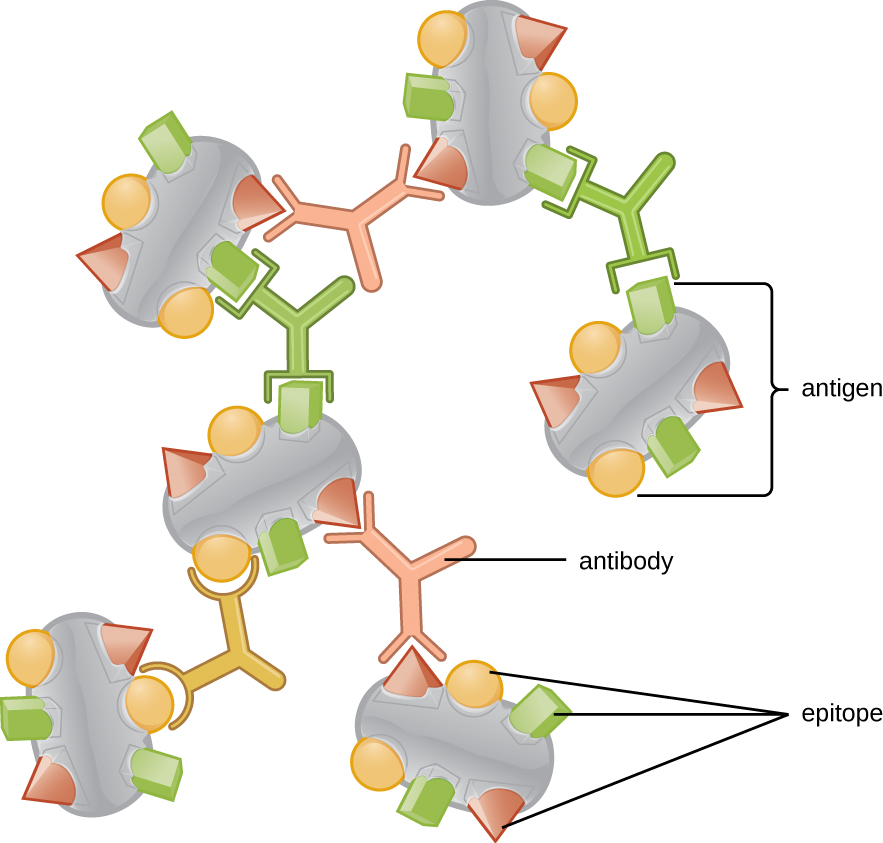
Одна з причин того, що тривимірна складність антигенів настільки важлива, полягає в тому, що антитіла та Т-клітини не розпізнають і взаємодіють з цілим антигеном, але з меншими оголеними ділянками на поверхні антигенів, які називаються епітопами. Один антиген може володіти декількома різними епітопами (рис.\(\PageIndex{2}\)), а різні антитіла можуть зв'язуватися з різними епітопами на одному антигені (рис.\(\PageIndex{3}\)). Наприклад, бактеріальний джгутик - це велика складна білкова структура, яка може володіти сотнями або навіть тисячами епітопів з унікальними тривимірними структурами. Більш того, джгутики різних видів бактерій (або навіть штамів одного виду) містять унікальні епітопи, які можуть бути пов'язані тільки специфічними антитілами.
Розмір антигену є ще одним важливим фактором його антигенного потенціалу. У той час як великі антигенні структури, такі як джгутики, мають кілька епітопів, деякі молекули занадто малі, щоб бути антигенними самі по собі. Такі молекули, звані гаптенами, по суті є вільними епітопами, які не входять до складу складної тривимірної структури більшого антигену. Щоб гаптен став антигенним, він повинен спочатку прикріпитися до більшої молекули носія (як правило, білка) для отримання кон'югатного антигену. Гаптен-специфічні антитіла, що виробляються у відповідь на кон'югатний антиген, потім здатні взаємодіяти з некон'югованими молекулами вільних гаптенів. Гаптени, як відомо, не пов'язані з будь-якими конкретними збудниками, але вони відповідають за деякі алергічні реакції. Наприклад, гаптен урушіол, молекула, виявлена в олії рослин, що викликають отруйний плющ, викликає імунну відповідь, яка може призвести до сильного висипу (званого контактним дерматитом). Аналогічно гаптен пеніцилін може викликати алергічні реакції на препарати класу пеніциліну.
Вправа\(\PageIndex{3}\)
- У чому різниця між антигеном і епітопом?
- Які фактори впливають на антигенний потенціал антигену?
- Чому гаптени, як правило, не антигенні, і як вони стають антигенними?
Антитіла
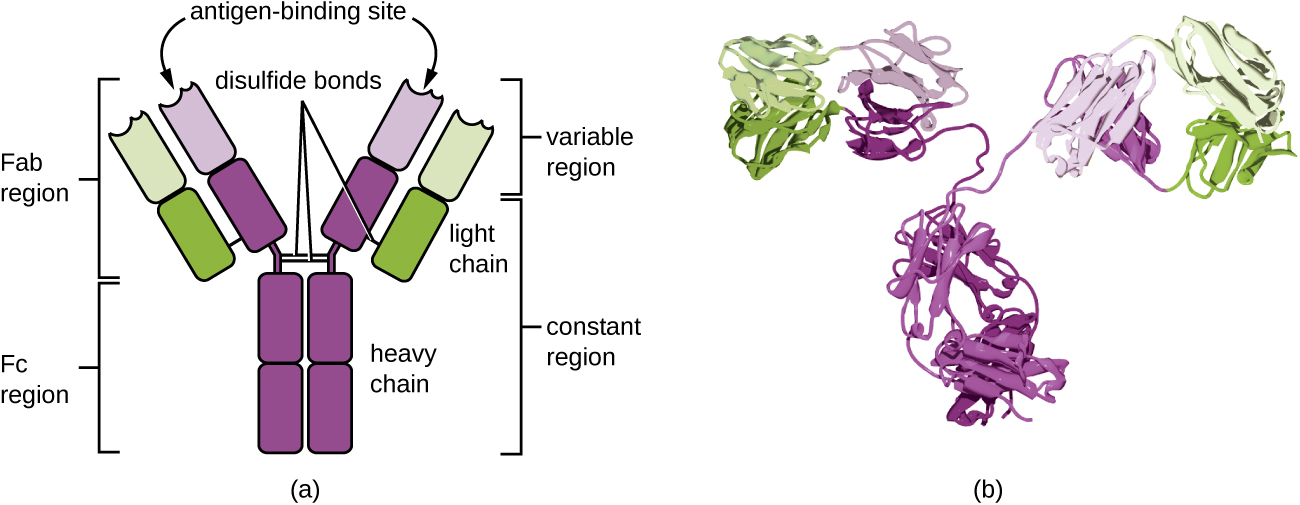
Антитіла (також звані імуноглобулінами) - це глікопротеїни, які присутні як в крові, так і в тканинних рідинях. Основна структура мономера антитіл складається з чотирьох білкових ланцюгів, скріплених між собою дисульфідними зв'язками (рис.\(\PageIndex{4}\)). Дисульфідний зв'язок - це ковалентний зв'язок між сульфгідрильними групами R, виявленими на двох амінокислотах цистеїну. Дві найбільші ланцюга ідентичні один одному і називаються важкими ланцюгами. Дві менші ланцюга також ідентичні один одному і називаються легкими ланцюгами. З'єднані між собою важкі і легкі ланцюги утворюють основну Y-подібну конструкцію.
Дві «руки» Y-подібної молекули антитіл відомі як область Fab, для «фрагмента зв'язування антигену». Дальнім кінцем області Fab є змінна область, яка служить місцем зв'язування антигену. Послідовність амінокислот у змінній області диктує тривимірну структуру, і, таким чином, специфічний тривимірний епітоп, з яким здатна зв'язуватися область Fab. Хоча епітопна специфічність областей Fab ідентична для кожного плеча однієї молекули антитіла, ця область демонструє високий ступінь мінливості між антитілами з різними специфіками епітопу. Зв'язування з областю Fab необхідно для нейтралізації патогенів, агглютинації або агрегації патогенів і антитілозалежної клітинної цитотоксичності.
Постійна область молекули антитіл включає стовбур Y і нижню частину кожного плеча Y. Стовбур Y також називається областю Fc, для «фрагмента кристалізації» і є місцем зв'язування фактора комплементу та зв'язування з фагоцитарними клітинами під час опосередкованої антитілами опсонізації.
Вправа\(\PageIndex{4}\)
Опишіть різні функції регіону Fab та регіону Fc.
Класи антитіл
Постійна область молекули антитіла визначає її клас, або ізотип. П'ять класів антитіл - IgG, IgM, IgA, IgD та IgE. Кожен клас має унікальні важкі ланцюги, позначені грецькими літерами γ, μ, α, δ і ε відповідно. Класи антитіл також демонструють важливі відмінності в достатку сироватки крові, розташування, місця дії тіла, функціональні ролі та розмір (рис.\(\PageIndex{5}\)).
IgG - це мономер, який на сьогоднішній день є найпоширенішим антитілом у крові людини, на який припадає близько 80% загального сироваткового антитіла. IgG ефективно проникає в тканинні простори, і є єдиним класом антитіл зі здатністю перетинати плацентарний бар'єр, забезпечуючи пасивний імунітет плоду, що розвивається під час вагітності. IgG також є найбільш універсальним класом антитіл з точки зору його ролі в захисті організму від патогенів.
IgM спочатку виробляється в мономерній мембранно-зв'язаній формі, яка служить антигензв'язуючим рецептором на В-клітині. Секретована форма IgM збирається в пентамер з п'ятьма мономерами IgM, пов'язаними між собою білковою структурою, яка називається ланцюгом J. Хоча розташування ланцюга J щодо областей Fc п'яти мономерів заважає IgM виконувати деякі функції IgG, десять доступних сайтів Fab, пов'язаних з пентамерним IgM, роблять його важливим антитілом в арсеналі захисних сил організму. IgM - це перше антитіло, що виробляється та секретується В-клітинами під час первинної та вторинної імунних реакцій, що робить специфічний для патогенів IgM цінним діагностичним маркером під час активних або недавніх інфекцій.
На IgA припадає близько 13% загального сироваткового антитіла, а секреторний IgA - найпоширеніший і рясний клас антитіл, що міститься в слизових виділеннях, які захищають слизові оболонки. IgA також можна знайти в інших виділеннях, таких як грудне молоко, сльози та слина. Секреторний IgA збирається в димерную форму з двома мономерами, з'єднаними білковою структурою, яка називається секреторним компонентом. Однією з важливих функцій секреторного IgA є затримка хвороботворних мікроорганізмів у слизу, щоб згодом їх можна було вивести з організму.
Подібно IgM, IgD - це мембранно-зв'язаний мономер, знайдений на поверхні В-клітин, де він служить антигензв'язуючим рецептором. Однак IgD не виділяється В-клітинами, а в сироватці крові виявляються лише слідові кількості. Ці слідові кількості, швидше за все, походять від деградації старих В-клітин і вивільнення молекул IgD з їх цитоплазматичних мембран.
IgE - найменш рясний клас антитіл у сироватці крові. Як і IgG, він виділяється як мономер, але його роль в адаптивному імунітеті обмежується антипаразитарним захистом. Область Fc IgE зв'язується з базофілами і огрядними клітинами. Потім область Fab пов'язаного IgE взаємодіє зі специфічними епітопами антигену, змушуючи клітини вивільняти потужні прозапальні медіатори. Запальна реакція, що виникає внаслідок активації огрядних клітин та базофілів, допомагає захищати від паразитів, але ця реакція також є центральною для алергічних реакцій (див. Захворювання імунної системи.)
Вправа\(\PageIndex{5}\)
- Яка частина молекули антитіла визначає її клас?
- Який клас антитіл бере участь в захисті від паразитів?
- Опишіть різницю в структурі між IgM і IgG.
Взаємодія антиген-антитіл
Різні класи антитіл відіграють важливу роль у захисті організму від патогенів. Ці функції включають нейтралізацію патогенів, опсонізацію для фагоцитозу, аглютинацію, активацію комплементу та цитотоксичність, опосередковану клітиною, залежну від антитіл. Для більшості цих функцій антитіла також забезпечують важливий зв'язок між адаптивним специфічним імунітетом і вродженим неспецифічним імунітетом.
Нейтралізація передбачає зв'язування певних антитіл (IgG, IgM або IgA) з епітопами на поверхні хвороботворних мікроорганізмів або токсинів, перешкоджаючи їх приєднанню до клітин. Наприклад, секреторний IgA може зв'язуватися зі специфічними патогенами і блокувати початкове приєднання до клітин слизової оболонки кишечника. Так само специфічні антитіла можуть зв'язуватися з певними токсинами, блокуючи їх від приєднання до клітин-мішеней і тим самим нейтралізуючи їх токсичну дію Віруси можна нейтралізувати і запобігти зараженню клітини тим же механізмом (рис.\(\PageIndex{6}\)).
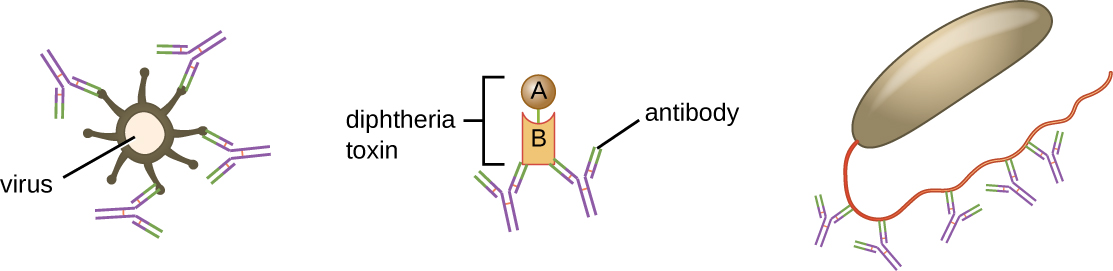
Як описано в Chemical Defences, опсонізація - це покриття збудника з молекулами, такими як фактори комплементу, С-реактивний білок та сироватковий амілоїд А, щоб сприяти зв'язуванню фагоцитів для полегшення фагоцитозу. Антитіла IgG також служать відмінними опсонінами, зв'язуючи їх Fab сайти зі специфічними епітопами на поверхні патогенів. Фагоцитарні клітини, такі як макрофаги, дендритні клітини та нейтрофіли, мають рецептори на своїх поверхнях, які розпізнають та зв'язуються з частиною молекул IgG Fc; таким чином, IgG допомагає таким фагоцитам приєднуватися та поглинати збудників, які вони зв'язали (Рис.\(\PageIndex{7}\)).
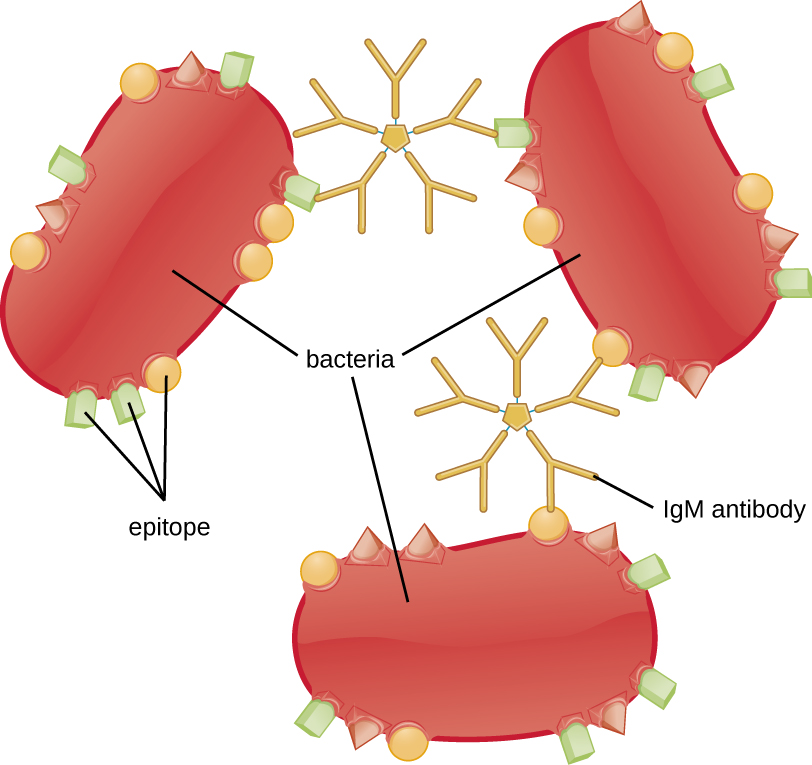
Аглютинація або агрегація передбачає зшивання патогенів антитілами для створення великих агрегатів (рис.\(\PageIndex{8}\)). IgG має два сайти зв'язування антигенів Fab, які можуть зв'язуватися з двома окремими клітинами збудника, комкуючи їх разом. Коли задіяні множинні антитіла IgG, можуть розвиватися великі агрегати; ці агрегати легше ниркам і селезінці фільтруються з крові і легше для фагоцитів для руйнування. Пентамерна структура IgMзабезпечує десять сайтів зв'язування Fab на молекулу, що робить її найбільш ефективним антитілом для аглютинації.
Ще одна важлива функція антитіл - активація каскаду комплементу. Як обговорювалося в попередньому розділі, система комплементу є важливим компонентом вроджених захисних сил, сприяючи запальній реакції, набору фагоцитів до місця інфекції, посилюючи фагоцитоз шляхом опсонізації та вбиваючи грамнегативні бактеріальні патогени за допомогою комплексу мембранної атаки (MAC) ). Активація комплементу може відбуватися трьома різними шляхами (див. Рис. 17.2.2), але найбільш ефективним є класичний шлях, який вимагає початкового зв'язування антитіл IgG або IgM з поверхнею клітини-збудника, що дозволяє рекрутувати і активувати комплекс С1.
Ще однією важливою функцією антитіл є антитіло-залежна клітинна цитотоксичність (ADCC), яка посилює вбивство збудників, які занадто великі, щоб бути фагоцитозованими. Цей процес найкраще характеризується для природних клітин-кілерів (NK-клітин), як показано на малюнку\(\PageIndex{9}\), але він також може залучати макрофаги та еозинофіли. ADCC виникає, коли Fab область антитіла IgG зв'язується з великим збудником; Fc-рецептори на ефекторних клітинах-клітинах( наприклад, NK-клітини) потім зв'язуються з областю Fc антитіла, приводячи їх у безпосередній близькості з цільовим збудником. Потім ефекторна клітина виділяє потужні цитотоксини (наприклад, перфорін та гранзими), які вбивають збудника.
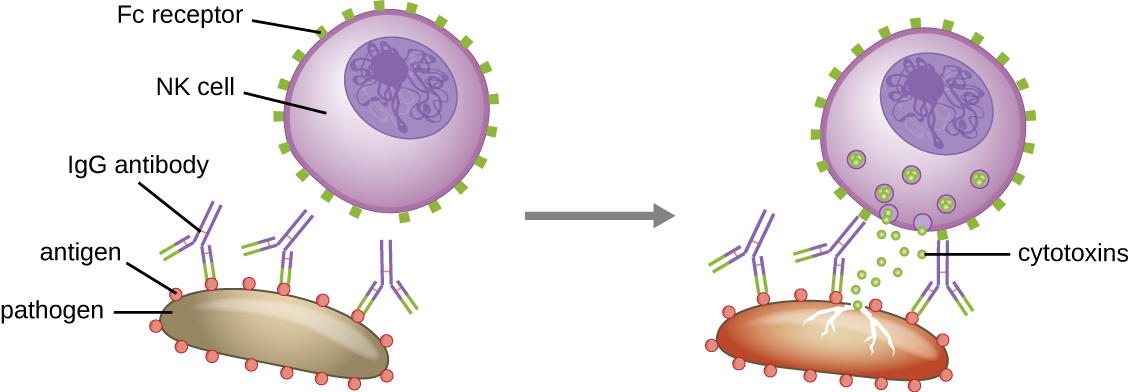
Вправа\(\PageIndex{6}\)
- Де зазвичай знаходиться IgA?
- Який клас антитіл перетинає плаценту, забезпечуючи захист плоду?
- Порівняйте механізми опсонізації та антитіло-залежної клітинно-опосередкованої цитотоксичності.
Ключові поняття та резюме
- Адаптивний імунітет - це придбаний захист від чужорідних збудників, яка характеризується специфічністю і пам'яттю. Перше вплив антигену стимулює первинну реакцію, а наступні впливу стимулюють більш швидку і сильну вторинну реакцію.
- Адаптивний імунітет - це подвійна система, що включає гуморальний імунітет (антитіла, що виробляються В-клітинами) і клітинний імунітет (Т-клітини, спрямовані проти внутрішньоклітинних збудників).
- Антигени, також звані імуногенами, - це молекули, які активують адаптивний імунітет. Одиночний антиген має менші епітопи, кожен з яких здатний індукувати специфічну адаптивну імунну відповідь.
- Здатність антигену стимулювати імунну відповідь залежить від декількох факторів, включаючи його молекулярний клас, молекулярну складність та розмір.
- Антитіла (імуноглобуліни) - це Y-подібні глікопротеїни з двома сайтами Fab для зв'язування антигенів та частиною Fc, що беруть участь у активації комплементу та опсонізації.
- П'ять класів антитіл - IgM, IgG, IgA, IgE та IgD, кожен з яких відрізняється розміром, розташуванням, розташуванням в організмі та функцією. П'ять первинних функцій антитіл - це нейтралізація, опсонізація, аглютинація, активація комплементу та цитотоксичність, опосередкована клітинами (ADCC), залежна від антитіл.