5: Бактерії - внутрішні компоненти
- Page ID
- 6631
Ми вже розглянули основні внутрішні компоненти, виявлені у всіх бактеріях, а саме цитоплазмі, нуклеоїді та рибосомах. Пам'ятайте, що бактерії, як правило, вважають, що не вистачає органел, тих біліпідних мембранно-зв'язаних відсіків, настільки поширених в еукаріотичних клітині (хоча деякі вчені стверджують, що бактерії мають структури, які можна вважати простими органелами). Але бактерії можуть бути більш складними, з безліччю додаткових внутрішніх компонентів, які можуть сприяти їх можливостям. Більшість цих компонентів є цитоплазматичними, але деякі з них периплазматичні, розташовані в просторі між цитоплазматичною і зовнішньою мембраною у грамнегативних бактерій.
Цитоскелет
Спочатку вважалося, що бактерії не мають цитоскелета, значного компонента еукаріотичних клітин. Однак за останні 20 років вчені виявили бактеріальні нитки, виготовлені з білків, які є аналогами цитоскелетних білків, виявлених у еукаріотів. Також було визначено, що бактеріальний цитоскелет відіграє важливу роль у формі клітин, поділі клітин та цілісності клітинної стінки.
ФЦЗ
FtSz, гомологічний до еукаріотичного білка тубуліну, утворює кільцеву структуру в середині клітини під час поділу клітин, залучаючи інші білки до області, щоб побудувати перегородку, яка врешті-решт відокремить дві отримані дочірні клітини.
МРеБ
MReb, гомологічний актину еукаріотичного білка, міститься в бацилах і спіралеподібних бактеріях і відіграє важливу роль у формуванні форми клітин. MReb передбачає спіральну конфігурацію, що працює на довжині клітини, і диктує діяльність пептидоглікан-синтезу машин, забезпечуючи несферичну форму.
Півмісяць
Кресцентин, гомологічний еукаріотичним білкам ламіну і кератину, міститься у спіралеподібних бактерій з єдиною кривою. Білок збирається уздовж у внутрішній кривизні клітини, згинаючи клітку в її кінцеву форму.
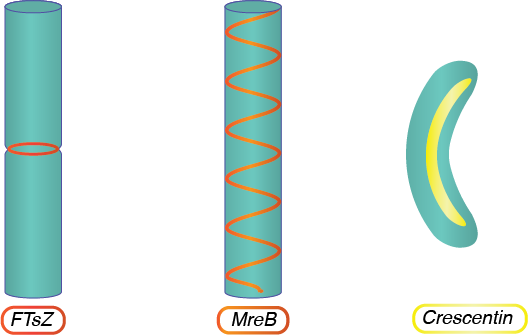
Цитоскелетні структури.
Включення
Бактеріальні включення, як правило, визначаються як виразна структура, розташована або всередині цитоплазми, або периплазми клітини. Вони можуть варіюватися за складністю, від простої компіляції хімічних речовин, таких як кристали, до досить складних структур, які починають конкурувати з еукаріотичними органелами, в комплекті з мембранним зовнішнім шаром. Їх роль часто полягає в зберіганні компонентів як метаболічних резервів для клітини, коли речовина виявляється в надлишку, але вони також можуть відігравати певну роль у рухливості та метаболічних функціях.
Зберігання вуглецю
Вуглець є найпоширенішою речовиною, яку зберігає клітина, оскільки всі клітини на основі вуглецю. Крім того, вуглецеві сполуки часто можуть швидко розщеплюватися клітиною, тому вони також можуть служити джерелами енергії. Одним з найпростіших і поширених включень для зберігання вуглецю є глікоген, в якому одиниці глюкози пов'язані між собою в багаторозгалуженій полісахаридній структурі.
Інший поширений спосіб зберігання вуглецю бактерій - це у вигляді полі-β-гідроксибутирату (PHB), гранули, яка утворюється, коли одиниці β-гідроксимасляної кислоти об'єднуються разом. Цей ліпідний за складом дуже пластичний, що змушує деяких вчених досліджувати можливість використання їх як біорозкладається пластика. Гранули PHB насправді мають оболонку, що складається як з білка, так і з невеликої кількості фосфоліпідів. І глікоген, і PHB утворюються, коли є надлишок вуглецю, а потім розщеплюються клітиною пізніше як для вуглецю, так і для енергії.
Неорганічне зберігання
Часто бактерії потребують чогось іншого, крім вуглецю, або для синтезу клітинних компонентів, або в якості альтернативного енергетичного запасу. Гранули поліфосфату дозволяють накопичувати неорганічний фосфат (PO43-), де фосфат може бути використаний для виготовлення нуклеїнової кислоти (пам'ятаєте цукрово-фосфатну кістку?) або АТФ (аденозин трифосфат, звичайно).
Інші клітини потребують сірки як джерело електронів для їх метаболізму і будуть зберігати надлишок сірки у вигляді сірчаних глобул, що призводить до того, коли клітина окислює сірководень (H2S) до елементарної сірки (S0), що призводить до утворення заломлюючих включень.
Функції, що не зберігаються
Бувають випадки, коли бактерії потрібно робити щось, крім простого зберігання органічних або неорганічних сполук для використання в метаболізмі, і є включення, які допомагають у цих функціях, що не зберігаються. Одним з таких прикладів є газові вакуолі, які використовуються кліткою для контролю плавучості в товщі води, забезпечуючи клітці певний контроль над тим, де вона знаходиться в навколишньому середовищі. Це обмежена форма моторики, тільки на вертикальній осі. Газові вакуолі складаються з конгломерацій газових бульбашок, циліндричних структур, які є як порожнистими, так і жорсткими. Газові бульбашки вільно проникні для всіх типів газів шляхом пасивної дифузії і можуть бути швидко побудовані або згорнуті, якщо це необхідно клітині для підняття або спуску.
Магнітосоми - це включення, що містять довгі ланцюги магнетиту (Fe3O4), які використовуються клітиною як компас в геомагнітних полах, для орієнтації в їх середовищі. Магнітотаксичні бактерії, як правило, мікроаерофільні, віддаючи перевагу середовищу з нижчим рівнем кисню, ніж атмосфера. Магенетосома дозволяє клітинам знайти оптимальну глибину для їх росту. Магенетосоми мають справжній ліпідний бішар, що нагадує еукаріотичні органели, але насправді це інвагінація плазматичної мембрани клітини, яка була модифікована специфічними білками.
Мікровідсіки
Бактеріальні мікрокомати (БМК) унікальні від інших включень в силу своєї структури і функціональності. Вони мають ікосаедричну форму і складаються з білкової оболонки, що складається з різних білків сімейства BMC. Хоча їх точна роль змінюється, всі вони беруть участь у функціях, що виходять за рамки простого зберігання речовин. Ці відсіки забезпечують як місце розташування, так і речовини (як правило, ферменти), необхідні для конкретної метаболічної діяльності.
Найкращим вивченим прикладом BMC є карбоксисома, яка міститься у багатьох бактеріях, що фіксують CO2. Карбоксисоми містять фермент рибулоза-1,5-бісфосфаткарбоксилазу (на щастя, він також відомий як RubiSCo), який відіграє вирішальну роль у перетворенні CO2 в цукор. Карбоксисома також відіграє певну роль у концентрації CO2, забезпечуючи тим самим, що компоненти, необхідні для фіксації CO2, знаходяться в одному місці одночасно.
Анаммоксома
Анамоксосома - це великий мембранно-зв'язаний відсік, виявлений у бактеріальних клітинам, здатних проводити анемоксну реакцію (аеробне окислення монію), де амоній (NH4+) та нітрит (NO2-) перетворюються на газ азоту (N2). Процес виконується як спосіб отримання клітиною енергії, використовуючи амоній як донор електронів і нітрит як прийом електронів, з отриманням газу азоту. Ця хімічна конверсія азоту має важливе значення для азотного циклу.
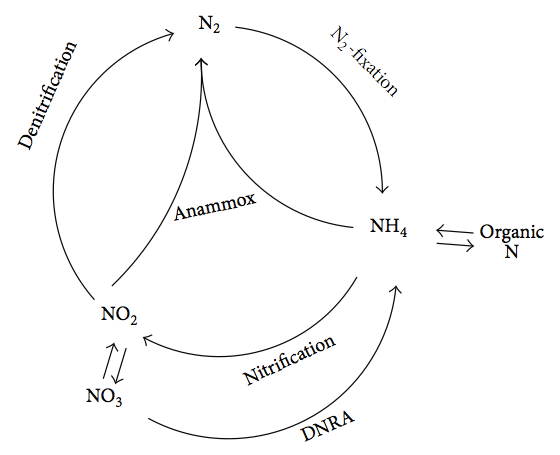
Цикл азоту. Шу Цін Ні та Цзянь Чжан [CC BY 3.0], через Вікісховище
хлоросом
Хлоросома, виявлена в деяких фототрофних бактеріях, є високоефективною структурою для захоплення низької інтенсивності світла. Вистилаючи внутрішній периметр клітинної мембрани, кожна хлоросома може містити до 250 000 молекул бактеріохлорофілу, розташованих в щільних масивах. Зібране світло передається реакційним центрам в клітинній мембрані, дозволяючи перетворюватися з світлової енергії в хімічну енергію у вигляді АТФ. Хлоросома обмежена ліпідним моношаром.
Плазміда
Плазміда - це позахромосомний шматок ДНК, який мають деякі бактерії, крім генетичного матеріалу, знайденого в нуклеоїді. Він складається з дволанцюгової ДНК і, як правило, кругової, хоча були знайдені лінійні плазміди. Плазміди описуються як «несуттєві» для клітини, де клітина може нормально функціонувати за їх відсутності. Але хоча плазміди мають лише кілька генів, вони можуть надати клітині важливі можливості, такі як резистентність до антибіотиків. Плазміди розмножуються незалежно від клітини і можуть бути втрачені (відомі як затвердіння) спонтанно або внаслідок впливу несприятливих умов, таких як УФ-світло, тимінове голодування або зростання вище оптимальних умов. Деякі плазміди, відомі як епізоми, можуть бути інтегровані в клітинну хромосому, де гени будуть репліковані під час поділу клітин.
Ендоспор
Потім є ендоспора, диво бактеріальної інженерії. Це знаходиться під заголовком «бактеріальні внутрішні компоненти», але важливо зазначити, що ендоспора не є внутрішньою або зовнішньою структурою стільки, скільки перетворенням клітини в альтернативну форму. Клітини починаються як вегетативна клітина, роблячи все, що повинна робити клітина (метаболізм, розмноження, косіння газону...). Якщо вони потрапляють у ворожі умови (десикація, висока спека, розлючений сусід...) і вони мають здатність, вони можуть перетворитися з вегетативної клітини в ендоспору. Ендоспора фактично формується всередині вегетативної клітини (хіба це не робить її внутрішньою структурою?) а потім вегетативна клітина лізує, вивільняючи ендоспору (чи робить це зовнішньою структурою?).
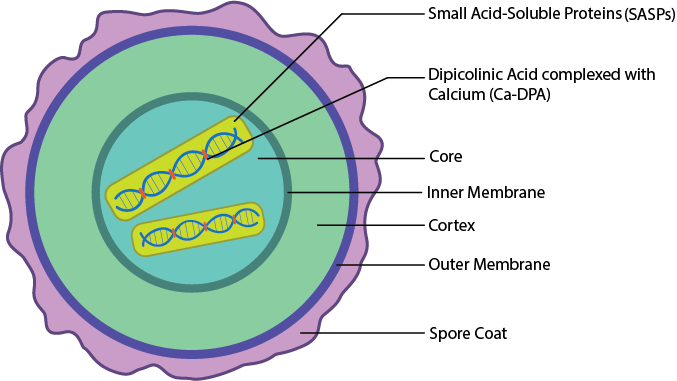
Ендоспорові шари.
Ендоспори утворюються лише кількома грамами позитивних пологів і забезпечують клітині стійкість до найрізноманітніших суворих умов, таких як голодування, перепади температури, вплив сушіння, ультрафіолетового світла, хімічних речовин, ферментів та випромінювання. Хоча вегетативна клітина є активною формою для бактеріальних клітин (вирощування, метаболізму тощо), ендоспора може розглядатися як спляча форма клітини. Вона дозволяє вижити несприятливі умови, але не дає клітині рости або розмножуватися.
Структура
Для того, щоб бути настільки неймовірно стійким до такої кількості різних речовин і умов навколишнього середовища, необхідно багато різних шарів. Ендоспора бактерій має безліч різних шарів, починаючи з ядра в центрі. Ядро - це розташування нуклеоїдів, рибосом і цитоплазми клітини, в надзвичайно зневодненому вигляді. Зазвичай він містить лише 25% води, що міститься у вегетативній клітині, підвищуючи жаростійкість. ДНК додатково захищена наявністю дрібних кислоторозчинних білків (SASP), які стабілізують ДНК і захищають її від деградації. Стабілізація ДНК підвищується наявністю дипіколінової кислоти, комплексної з кальцієм (Ca-dPA), яка вставляється між основами ДНК. Ядро загорнуте у внутрішню мембрану, яка забезпечує бар'єр проникності хімічних речовин, який потім оточений корою, товстим шаром, що складається з пептидоглікану з меншим зшиванням, ніж знаходиться у вегетативній клітині. Кора загорнута зовнішньою оболонкою. Нарешті, кілька спорових покриттів, виготовлених з білка, які забезпечують захист від стресу навколишнього середовища, таких як хімічні речовини та ферменти.
Sporulation: перетворення з вегетативної клітини в ендоспору
Sporulation, перетворення вегетативної клітини в високозахисну ендоспору, зазвичай відбувається, коли виживання клітини певним чином загрожує. Фактичний процес дуже складний і зазвичай займає кілька годин до завершення. Спочатку спорулюючі клітини реплікують свою ДНК, ніби вона ось-ось піддається поділу клітин. Перегородка утворюється асиметрично, секвеструючи одну копію хромосоми на одному кінці клітини (званої передньою спорою). Відбувається синтез ендоспороспецифічних речовин, що змінюють передміхурову спору і призводять до розвитку шарів, специфічних для ендоспори, а також зневоднення. Зрештою «материнська клітина» - це лізи, що дозволяють вивільнити зрілу ендоспору в навколишнє середовище.
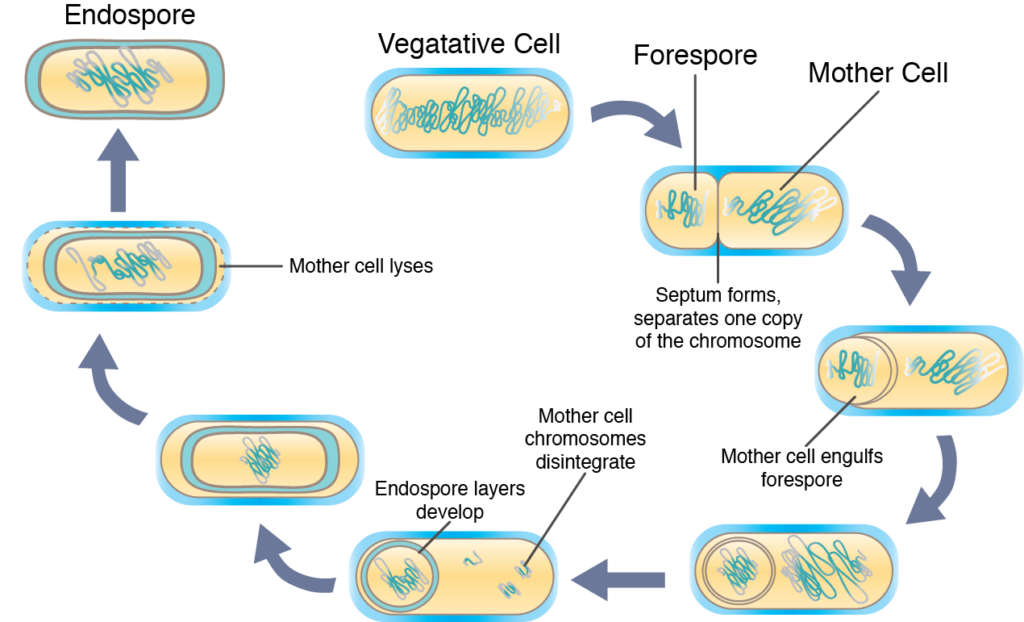
Спорокуляція.
Перетворення з ендоспори в вегетативну клітину
Ендоспора залишається в стані спокою, поки умови навколишнього середовища не покращаться, викликаючи хімічну зміну, яка ініціює експресію генів. Існує три різних етапи перетворення з ендоспори в метаболічно активні вегетативні клітини: 1) активація, етап підготовки, який може бути ініційований застосуванням тепла; 2) проростання, коли ендоспора стає метаболічно активною і починає приймати на себе вода; 3) наріст, коли вегетативна клітина повністю виходить з ендоспорової оболонки.
Ключові слова
цитоскелет, FtSz, тубулін, mReB, актин, кресентин, ламін, кератин, включення, глікоген, полі-β-гідроксибутират (PHB), поліфосфатна гранула, сірчана глобула, газова вакуоля, газова везикула, магнітосома, мікроаерофільна, мікровідсік, бактеріальні мікровідділення (BMC), карбоксисома, рибулоза 1,5-біфосфат карбоксилаза, RubiSCo, анамоксома, анемоксома реакція, хлоросома, плазміда, затвердіння, епісома, ендоспора, вегетативна клітина, ядро, малі кислоторозчинні білки (SASP), дипіколінова кислота, Ca-DPA, внутрішня мембрана, кора, зовнішня мембрана, спорова оболонка, спороутворення, передмова спора, активація, проростання, розростання.
Основні питання/цілі
- Які ролі і склад бактеріального цитоскелета? Чим він відрізняється від еукаріотичного цитоскелета? Які специфічні бактеріальні білки цитоскелета і які подробиці відомі про кожного?
- Для чого призначаються включення, виявлені в бактеріях? Які їх характеристики?
- Які конкретні приклади накопичувальних включень містяться в бактеріях? Вміти описати кожен тип з точки зору структури і призначення.
- Які ще включення містяться в бактеріях? Вміти описати кожен тип з точки зору структури і призначення.
- Чим мікрокупе відрізняються від включень? Які конкретні приклади? Який склад і призначення?
- Що таке анаммоксосоми? Який їх склад і призначення?
- Що таке плазміди і якими характеристиками вони володіють? Що таке епізоми? Що таке лікування і що його викликає?
- Що таке бактеріальні ендоспори? Яке їх призначення? Якими характеристиками вони володіють? Які існують різні шари ендоспори і яку роль відіграє кожен шар?
Дослідницькі питання (НЕОБОВ'ЯЗКОВО)
- Які бактеріальні структури можуть бути корисні вченим у вирішенні соціальних проблем?