8.4: Пост-транскрипційна обробка РНК
- Page ID
- 3683

Перший з пост-транскрипційних подій 5' кінець обмеження. Після того, як 5' кінець зароджується РНК поширюється вільно від RNAP II приблизно 20-30 nt, він готовий до обмеження 7-метилгуанозинової структурою. Ця 5' «шапка» служить місцем розпізнавання для транспортування завершеної мРНК з ядра і в цитоплазму.
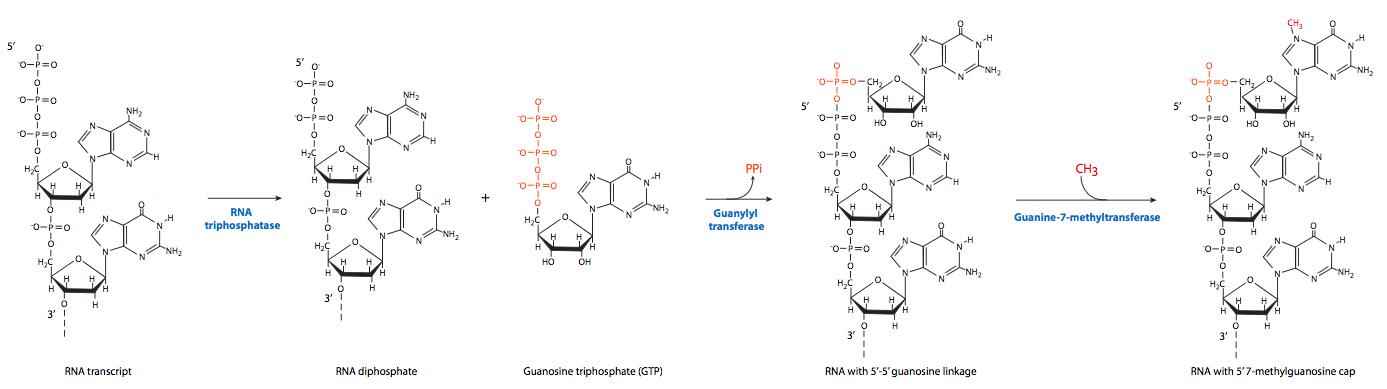
Процес насправді включає в себе три етапи. По-перше, РНК-трифосфатаза видаляє 5-кінцеву трифосфатну групу. Гуанілювання GTP каталізується шляхом укупорки ферменту, утворюючи незвичайну 5'-5' «зворотну» зв'язок між новим гуаніном і першим нуклеотидом стенограми РНК. Нарешті, гуанін-7-метилтрансфераза метилює знову приєднаний гуанін.
На протилежному кінці РНК, на вільному 3'-ОН, відбувається поліаденілювання. Як зазначалося раніше, ферментний комплекс, який приєднується до ділянки на Хвості CTD RNAP II, розщеплює частину 3-футового кінця поблизу послідовності розпізнавання AAUAAA, а потім послідовно додає велику кількість залишків аденіну. Полі (А) хвіст не потрібно для перекладу, але він впливає на стабільність транскриптів в цитоплазмі. Оскільки молекули мРНК довше залишаються в цитоплазмі, хвостик полі (А) поступово видаляється. Після того, як полі (А) хвіст зникне, мРНК незабаром буде зруйнована. Молекули мРНК з довшими хвостами полі (А), як правило, довше живуть в цитоплазмі, ніж ті, у кого коротші хвости, але в даний час немає доказів прямо пропорційного ефекту.
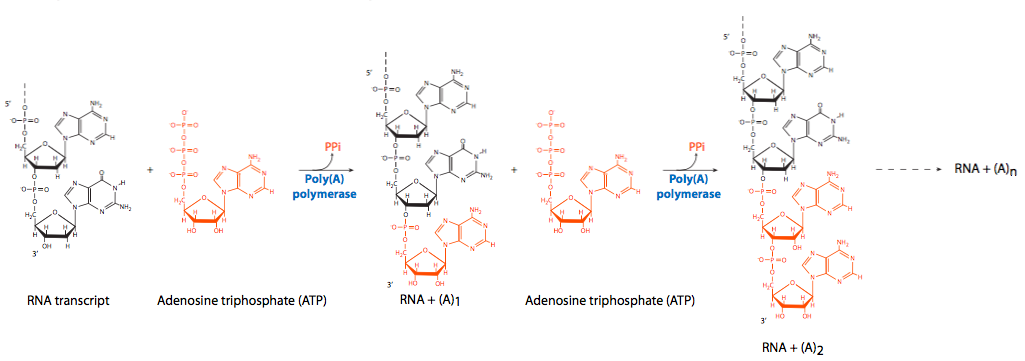
Хоча фермент, який розщеплює первинну розшифровку при підготовці до поліаденілювання, не був ідентифікований, були залучені два неферментативні фактори, збуджуюче названий фактор розщеплення I (CFI) та фактор розщеплення II (CFII). Серійне аденілювання походить від активності полі (А) полімерази (PAP) у поєднанні з CPSF (фактор розщеплення та поліаденілювання speci city), який зв'язується з РНК. Сам PAP має відносно погану спорідненість до РНК. Як і в інших полімеразах нуклеїнових кислот, він додає нові нуклеотиди на вільний 3'-ОН вже існуючого ланцюга. Для заохочення технологічності (безперервної полімеризації) полі-( А) зв'язуючий білок II (PABII) приєднується до поліаденілірующего комплексу і бере участь у контролі кінцевої довжини полі (А) хвоста. Слід зазначити, що PABII є ядерним білком і його не слід плутати з PABP (poly (A) зв'язуючим білком), який зв'язується з молекулами мРНК в цитоплазмі і відіграє певну роль у захисті їх від атаки нуклеази.
Третя і найскладніша модифікація новотранскрибованої еукаріотичної РНК - це зрощування. На відміну від прокаріотичної РНК, яка є безперервно трансльованою областю кодування відразу ж, коли вона виходить з РНК-полімерази, більшість еукаріотичних РНК мають перервані області кодування. Зрощування - це процес, за допомогою якого некодуючі області, відомі як інтрони, видаляються, а області кодування, відомі як екзони, з'єднуються разом. У деяких РНК це може відбуватися автономно, при цьому частина РНК виступає ферментативним каталізатором процесу. Це вимагає, щоб РНК мала специфічну вторинну та третинну структуру, приводячи два екзони близько один до одного, виводячи інтрон. Саме вивчення цього явища призвело до відкриття рибоцимів, які є ферментами, виготовленими з РНК.
До відкриття рибоцимів передбачалося, що лише ферменти можуть генеруватися лише з різноманітністю структур, можливих з амінокислотами в білках.
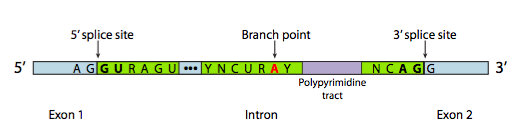
Однак у більшості випадків сплайсинг здійснюється багатосубодиничним білковим комплексом, відомим як сплацеосома. Незалежно від того, чи є це самозрощення або spliceosome, є три основні компоненти послідовності, необхідні для визначення інтрона, який буде зрощений (Рисунок\(\PageIndex{8}\)). Існує 5-дюймовий сайт зрощування з послідовністю консенсусу AG|GUAAGU. Існує 3' місце зрощування, яке починається з 11-нуклеотидного поліпіримідинового тракту, за яким слідує NCAG | G. І десь між ними є аденін гілкової точки, як правило, в послідовності YNCURAY (Y - піримідин, N - будь-який нуклеотид, R - пурин). Зрощування насправді є сукупністю двох послідовних реакцій переетерифікації, і вимагає фізичної близькості реактивних ділянок шляхом згинання та зациклювання РНК, або автономно, або навколо білкових факторів, відомих як SNRNP (виражені «snurps»). SNRNP є абревіатурою для малих ядерних рибонуклеопротеїнів. Вони містять як білок, так і невеликий компонент ядерної РНК (SNRNA); останній допомагає при розпізнаванні послідовності. Дослідження структури частини SNRNA цих сплацеосом SNRNP показує, що вони дуже схожі на форми, прийняті самим стенограмою РНК у випадках самозрощування. Маючи це на увазі, більша частина наступного опису сплайцеосом опосередкованого зрощування відбувається в самозрощування, а також.
Хоча SNRNP є основними компонентами сплайсеосоми, різні інші фактори зрощування також відіграють певну роль. Найбільш помітними є U2AF (U2-асоційований фактор, який зв'язується з поліпіримідиновим трактом, і SF1 (фактор сплайсингу 1, він же гілка-точка білка BPP), який зв'язується з послідовністю консенсусу поблизу точки розгалуження. Разом вони допомагають правильно розташувати СНРНП U2. Існує також безліч інших менш вивчених факторів сплайсингу з сімейства білка SR (C-термінальний мотив зв'язування серин-аргініну) та сімей HNRNP (гетерогенний ядерний рибонуклео-білок), які діють для набору первинних членів сплайсеосоми до їх належних місць.
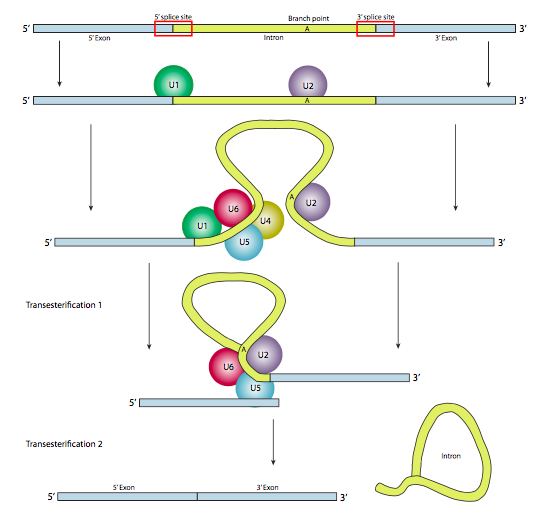
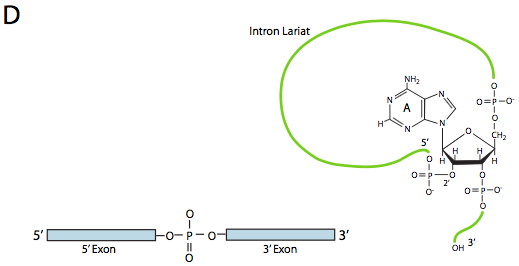
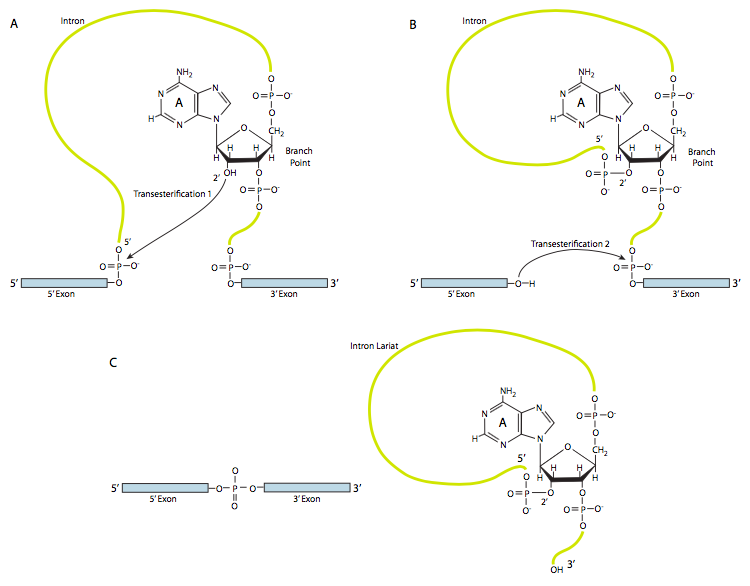
На першому кроці U1 SnRnP зв'язується з внутрішньою частиною 5-дюймового місця зрощування. Далі U2 SnRnP зв'язується з сайтом консенсусу навколо точки розгалуження, але важливо, що немає сполучення бази з самою гілкою A. Натомість через базове сполучення U2 з навколишньою послідовністю точка гілки А змушена випирати з решти РНК у цьому регіоні. U4, U5 та U6 приєднуються до сплайцеосоми разом, але в той час як U5 зв'язується з екзоном 5', а U6 витісняє U1 на місці сплайсингу 5', U4 лише тимчасово приєднаний, а також падає з сплайцеосоми перед першою реакцією переетерифікації. Як показано на малюнку, у цій реакції 2'-ОН точки розгалуження А нуклеофілічно атакує 5'-фосфат першого інтронного нуклеотиду, утворюючи структуру ларіату, в якій 5' кінець інтрона з'єднаний з точкою розгалуження через зв'язок 2',5'-фосфодіефіру. Це звільняє 5' ексон (і всю 5' половину РНК з цього приводу), але він зберігається в безпосередній близькості від 3' екзону (і решти РНК) U5, який приєднується до обох екзонів. Це дозволяє проходити другу переетерифікацію, в якій 3'- ОН першого екзону атакує 5' фосфат на початку другого екзону, тим самим одночасно розриваючи зв'язок між інтроном і другим екзоном, а також з'єднуючи два екзони через звичайний 3',5'-фосфодіефірний зв'язок . Таким чином, інтрон у формі ларіату звільняється і швидко деградує.
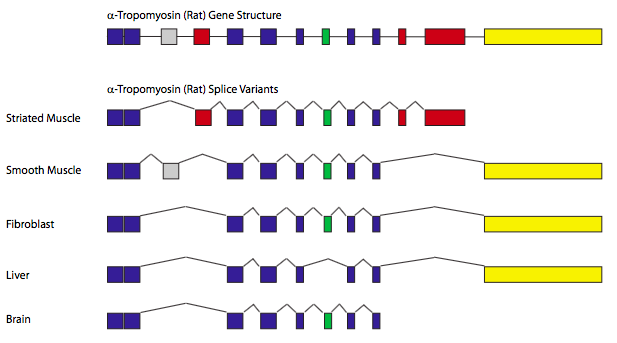
Зрощування - це ефективний (щодо розміру генома) спосіб генерувати різноманітність білка. При альтернативному зрощуванні деякі потенційні інтрони можуть бути зрощені за певних обставин, але залишаються як послідовність кодування за інших обставин. Нагадаємо, що місця зрощування розпізнаються шляхом сполучення з базою, і, отже, можуть бути сильніші та слабші місця зрощування залежно від того, наскільки вони близькі до консенсусу та додаткової послідовності на SNRNP. Тому ген з декількома потенційними інтронами може мати всі інтрони зрощені 80% часу, але інші 20% часу, можливо, лише один або два інтрони зрощуються. Додаючи мінливість, існують фактори зрощування, які можуть зв'язувати поблизу місць зрощування і можуть або зробити їх більш легко впізнаваними, або майже прихованими.
Класичним прикладом альтернативного сплайсингу є ген, що кодує α-тропоміозин (рис.\(\PageIndex{11}\)). Шляхом сплайсингу в/з різних комбінацій екзонів, один ген може генерувати сім різних білків, залежно від типу тканини. У цих випадках конкретні типи клітин або тканин містять специфічні комбінації факторів зрощування, а отже, контролюють розпізнавання конкретних місць зрощування, що призводить до різних моделей зрощування.
Хоча на цьому обговорення основних механізмів транскрипції завершується, наступна глава дійсно є продовженням цього: контроль експресії генів у найпростішій формі регулює розпізнавання послідовності промоторів РНК-полімеразою.