8.2: Прокаріотична транскрипція




- Last updated
- Oct 24, 2022
- Save as PDF

У кишкової палички, як і у інших прокаріотів, існує лише одна справжня РНК-полімераза (не враховуючи спеціальність РНК-полімерази, примази, яка робить короткі РНК-праймери для реплікації ДНК). Полімераза являє собою багатосубодиничний голофермент, що складається переважно з двох α субодиниць, β субодиниці, β' субодиниці, ω субодиниці та субодиниці σ. Субодиниці α в першу чергу структурні, що збирають голоензим і пов'язані з ними регуляторні фактори. β субодиниця містить полімеразну активність, яка каталізує синтез РНК, тоді як β' субодиниця використовується для неспецифічного зв'язування з ДНК. Субодиниця ω бере участь у складанні голоферменту, а також може відігравати роль у підтримці структурної цілісності РНК-полімерази. Нарешті, існує субодиниця σ, яка не залишається тісно пов'язаною з основним ферментом (αββ'ω), за винятком випадків, коли допомагає ініціювати транскрипцію, і використовується для розпізнавання промотора шляхом одночасного зменшення спорідненості RNAP до ДНК загалом, але збільшуючи спорідненість РНАП до конкретної ДНК послідовності промоутерів. Чому знижується спорідненість до неспецифічної ДНК? Коли РНАП не використовується, він не просто плаває в нуклеоплазмі: він досить щільно пов'язаний уздовж ДНК. Коли сигма пов'язана, знижена спорідненість дозволяє голоферменту RNAP рухатися вздовж ДНК та сканувати послідовності промоторів. Існує кілька ізоформ σсубодиниці (наприклад, сигма-70, згаданої вище), кожна з яких розпізнає різні послідовності промоторів. Всі ізоформи виконують одну і ту ж основну функцію правильного розташування РНПД до початку гена, і всі ізоформи залишаються прикріпленими до голоферменту лише для цієї перехідної мети, після чого вони вивільняються (зазвичай після транскрибування близько десяти нуклеотидів).
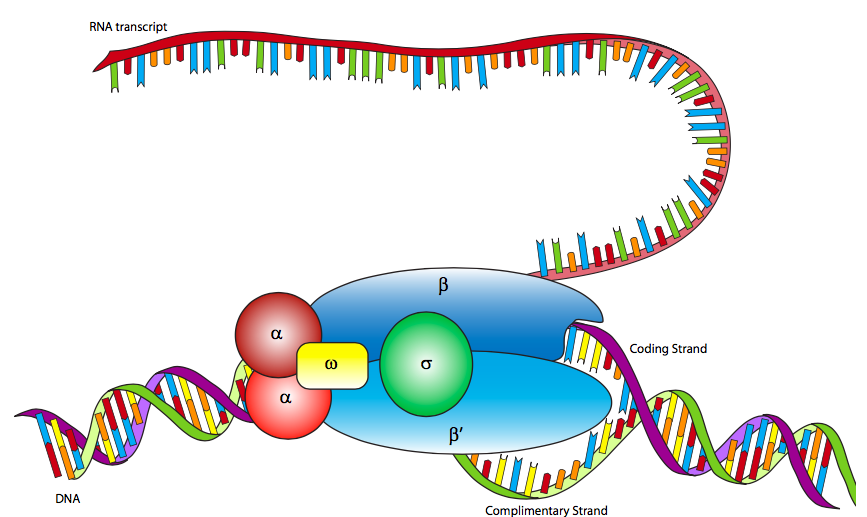
Хоча РНК-полімераза була виявлена в 1960 році, RNAP кишкової палички ще не був успішно відображений рентгенівською кристалографією. Однак він дуже схожий на RNAP виду археї, Thermophilus aquaticus, який є високостабільним (= легше кристалізується) і для якого з'ясована рентгенівська кристалографічна структура. Дані структури Taq RNAP та електронно-мікроскопічних аналізів E. coli RNAP виробляють голофермент у формі лобстера у формі кігтів. Внутрішня поверхня кігтя вистелена позитивно зарядженими амінокислотами, які можуть взаємодіяти з негативно зарядженою ДНК, і коли голофермент зв'язує сигма-субодиницю, дві половини кігтя (утворені в основному бета- і бета-субодиницями) рухаються ближче один до одного, щоб взаємодіяти з ДНК.
Рифаміцин - це клас антибіотиків, які включають рифаміцин В, виготовлений бактеріями Streptomyces mediterranei (до речі, лише один з багатьох антибіотиків, отриманих з роду Streptomyces) та рифампіцином, його синтетичним двоюрідним братом. Вони працюють шляхом зв'язування всередині каналу ДНК-РНК поблизу активної ділянки РНК-полімерази, що стерильно перешкоджає приєднанню нуклеотидів до ланцюга РНК. Якщо організм не може транскрибувати РНК, він також не може використовувати РНК, щоб зробити ферменти та інші білки, необхідні для життя, і помирає. Місце зв'язування рифаміцину дуже зберігається у більшості прокаріотів, але не в еукаріотів, тому антибіотик вбиває бактерії спеціально з невеликою ймовірністю завдати шкоди еукаріотів.
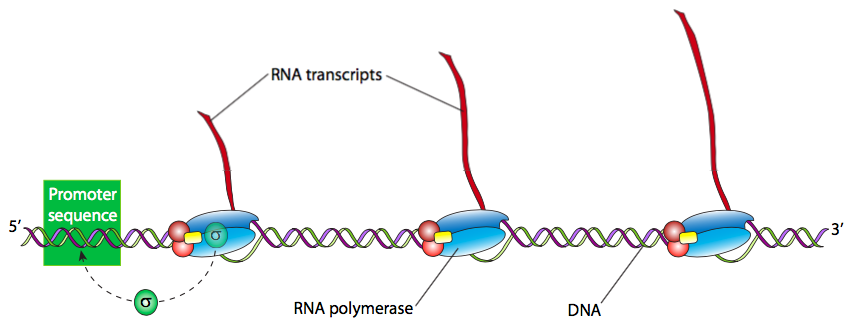
Після того, як голофермент розпізнає і щільно зв'язався з ДНК на сайті промотора, наступним кроком є «розплавлення» ДНК (розрив H-зв'язків і відокремлення ниток подвійної спіралі) в цій області, щоб RNAP міг протікати вниз за течією, прочитати шаблон ланцюга ДНК та виробляти нову РНК. Часто багато стенограми РНК гена необхідні для отримання великої кількості активних білків за короткий проміжок часу. Тому високотранскрипційно активні гени часто мають кілька РНК-полімераз, які читають їх один за одним. Як правило, РНК-полімеразі потрібно обробити лише близько 15 нуклеотидів, перш ніж з'явиться місце для іншого RNAP, може зв'язати промотор і почати іншу розшифровку.
Поділ ниток є енергетично важким процесом через міцність комбінованих H-зв'язків між пасмами, і часто RNAP може зробити кілька короткочасних абортивних спроб, перш ніж нарешті захопитися відкрити подвійну спіраль довго і досить далеко, щоб дозволити RNAP стабілізувати і транскрибувати безперервно зупинка сайту.
Фаза подовження транскрипції протікає в напрямку від 5 до 3', тобто нові нуклеотиди додаються до 3'-OH зростаючої пасма. Подовження - це стохастичний процес, при якому один з рясних вільно плаваючих рибонуклеотидів опускається в активну ділянку РНПД навпроти шаблону ДНК. Якщо це правильний нуклеотид (доповнює шаблон), то H-зв'язки тимчасово утворюватимуться, стабілізуючи новий нуклеотид на місці досить довго, щоб РНП каталізував утворення фосфодіефірного зв'язку між 3'-OH РНК-в-прогрес і 5'-фосфатом нуклеотиду. Однак, якщо це неправильний нуклеотид, належні Н-зв'язки не утворюються, і нуклеотид зазвичай дисоціює від активної ділянки до того, як RNAP матиме шанс зв'язати його з зростаючою ниткою РНК. Очевидно, що це не досконала система, і насправді частота помилок при транскрипції досить висока - приблизно 1 з 10000 нуклеотидів. На щастя, клітина, як правило, дуже швидко виписує багато копій РНК з будь-якого даного гена (приблизно 80 нуклеотидів в секунду), більшість з яких або безпомилкові, або мають помилки, які не впливають на функцію білка кінцевого продукту. Крім того, на відміну від ДНК, в якій помилки реплікації переносяться від одного покоління клітин до іншого, РНК не є середовищем зберігання, і її минуща природа означає, що навіть мутації, які сильно впливають на функцію білка, впливають лише на кілька білків, перекладені з цієї однієї РНК, а не білки генеруються з інших РНК, виготовлених з того ж шаблонного гена, набагато менше наступних поколінь. Іншими словами, щоб неправильно привласнити фразу з фільму Фрикадельки, «Це просто не має значення».
Зрештою, РНК-полімераза досягає кінця гена і припиняє транскрибування. Місце закінчення зазвичай позначається послідовністю залишків аденіну (А) 4-10 на нитку шаблону, а деякі мають паліндромну область, багату G-C, яка утворює петлю шпильки трохи вище за течією ряду аденінів. У першому випадку вважається, що результуючий рядок пар основ A-U нестійкий і може призвести до того, що РНК і нова нитка РНК випадає з ДНК шаблону, в той же час структура шпильки може спричинити зупинку або паузу RNAP, і це також може призвести до її дисоціації від ДНК. Лише близько половини всіх сайтів припинення транскрипції позначені таким чином, а інші не мають значних петель шпильок або легко впізнаваних послідовностей, крім серії багатих на G-C областей. У цьому типі місця закінчення, ферментативний кофактор, rho, необхідний для припинення, і тому це відомо як rho-залежне припинення. Rho - це РНК-зв'язуючий білок з хеліказной активністю, тому постулюється, що він впливає на припинення, змушуючи нитку РНК від шаблону ДНК.