4.1: Вступ: Велика картина
- Page ID
- 3369

Ми вже почали вивчати генетику розвитку і і отримали смак складності генетичних взаємодій. Наприклад, ген дрозофіли Бікоїд транскрибується в мРНК в материнських клітині і закачується в ембріон. Там його експресія регулюється сигналами локалізації на мРНК, а також деградацією і локалізацією білка. Бікоїдний білок сам по собі діє як регулятор транскрипції і трансляції нижележащих генів. Кінцевим ефектом є ембріон з різними генами, вираженими по осі A/P.
Тепер ми зробимо крок назад і подивимося на більш широку картину генетики розвитку, з точки зору сигнальних шляхів. Сигнальний шлях дозволяє клітинам спілкуватися зі своїм зовнішнім середовищем. У біології розвитку, це, як правило, клітинно-клітинна взаємодія. Такі взаємодії неймовірно важливі, оскільки кожна клітина повинна слідувати своїй власній траєкторії розвитку в координації з усіма клітинами навколо неї. Наприклад, уявіть зростаючу бруньку кінцівки ссавців. Кожній клітині в нирці кінцівки потрібно знати, чи знаходиться вона на великому пальці або мізинчастої стороні кінцівки і наскільки вона близька до тіла. Мало того, він також повинен знати, як довго прогресував розвиток. Уявіть, якби остеогенна клітина почала формувати кістковий матрикс на початку розвитку кінцівки. Він не зазнав би достатньої кількості клітинних відділів, щоб зробити правильну кількість остеобластів, що призводить до занадто маленької кістки та тварини з неправильною кінцівкою.
Генетичний інструментарій
(Вступний PDF можна знайти тут: Повторне відкриття біології www.learner.org/курси/біологія/pdf/7_gendev.pdf)
Одним з основних висновків Evo-Devo є те, що організми мають загальний набір генів для побудови своїх дуже різноманітних тіл. Зокрема, тварини поділяють набір генів «плану тіла», які виконують подібні функції в розвитку. Змішування та узгодження різних моделей експресії генів у типах тканин та клітин, здається, є основним механізмом для генерації унікальних тіл, які ми бачимо в кожній тварині. Нові еволюційні особливості (такі як шипи, пір'я, патерні серця, щупальця тощо), здається, покладаються на повторне використання генів інструментарію в новий час та місця в розвитку (див. Нескінченні форми для більш докладної інформації). Нижче ми почнемо розглядати генетику, яка стоїть за цим генетичним інструментарієм - як ці гени взаємодіють один з одним?
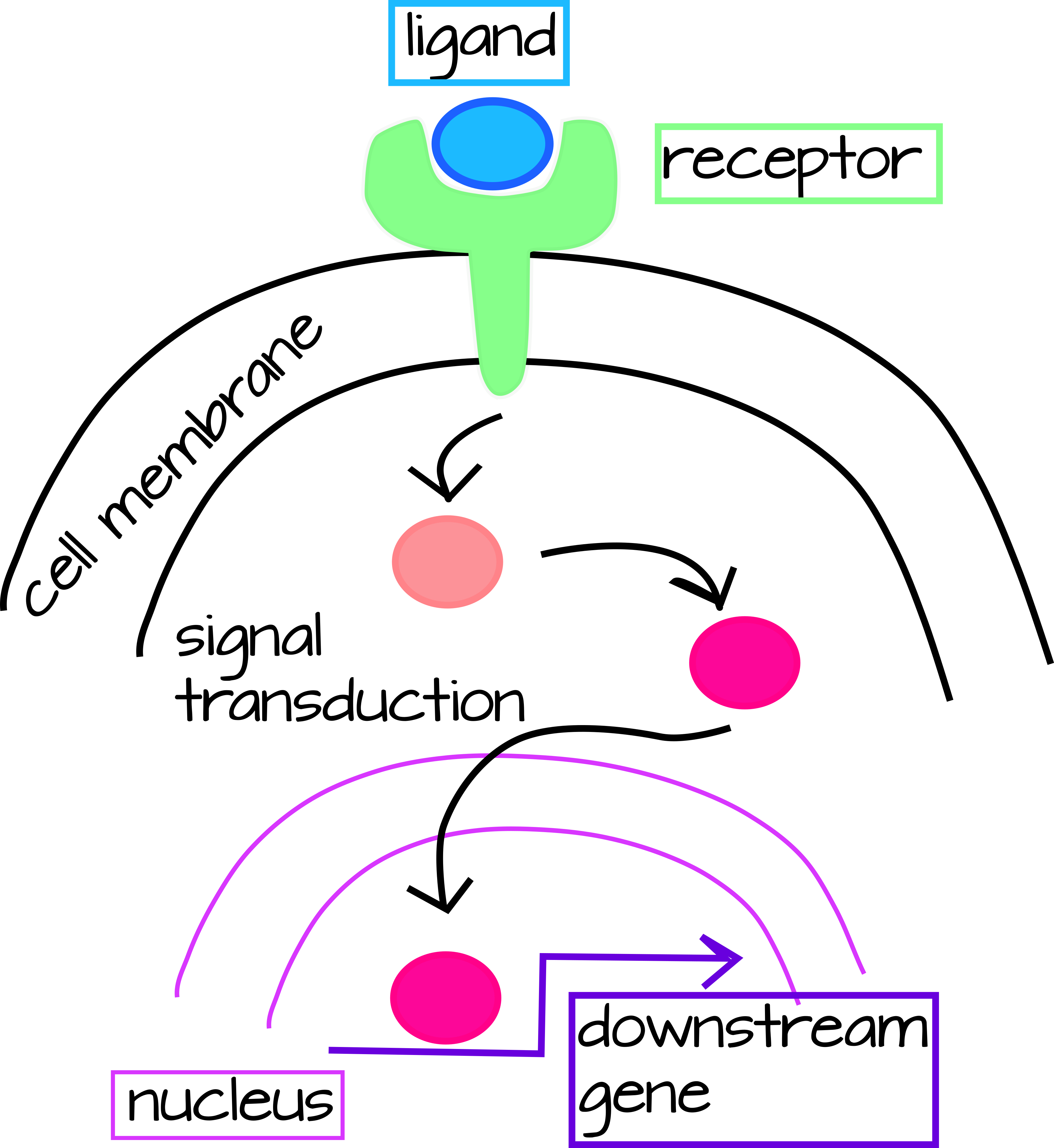
У Evo/Devo є два основні способи, якими ми можемо думати про генетичні взаємодії, але оскільки вони взаємопов'язані, ми розглянемо обидва одночасно. Одним з них є Шляхи трансдукції сигналу, які мають зовнішній сигнал (в біології розвитку це, як правило, секретований білок), який зазвичай приймається клітинно-мембранним рецептором, який експресують лише певні клітини. Коли ліганд (секретований білок) і рецептор зв'язуються, це запускає каскад білкових модифікацій, які призводять до транскрипційної активації нижчих генів (рис. 1).
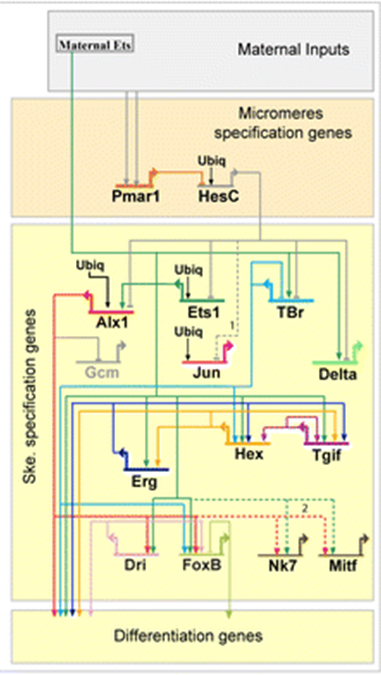
Другий спосіб Evo/Devo думає про сигналізацію - це генні регуляторні мережі (GRN). Сюди входить шлях трансдукції сигналу та його подальший вплив на експресію генів. Якщо ми збільшимо експресію одного гена, на які інші гени він впливає? GRN часто представлені у вигляді монтажних схем зі стрілками і смугами, що показують вплив білків на експресію генів, що знаходяться нижче за течією (рис. 2).
 |
.png) |
| Малюнок 1: Загальний шлях передачі сигналу. Позаклітинний ліганд зв'язується з мембрано-зв'язаним рецептором, запускаючи активацію каскаду трансдукції сигналу в цитоплазмі. Цей каскад може бути простим (як у випадку шляху Notch нижче) або дуже складним, що вимагає декількох кроків і типів введення. Останнім етапом трансдукційного каскаду є транслокація фактора цитоплазматичної транскрипції до ядра, де він активує або пригнічує транскрипцію гена нижче за течією. Редагований файл svg цього малюнка можна завантажити за адресою https://scholarlycommons.pacific.edu/open-images/18/ | Малюнок 2: Часткова схема регулювання генів мережі для специфікації скелетоутворюючої мезодерми у морського їжака, Strongylocentrotus purpuratus. Кожен ген представлений горизонтальною лінією, прикріпленою до стрілки такого ж унікального кольору. Горизонтальна лінія вказує на цис-регуляторну область гена, а стрілка вказує на транскрипцію області кодування мРНК. Лінії, що виходять з області кодування мРНК, вказують на те, як білковий продукт гена регулює інші нинішні гени. Входи в цис-регуляторні області білкових продуктів інших генів можуть бути активуючими (стрілками) або інгібуючими (бар). «Ubiq» - невідомі активатори, виражені у всіх клітині. Малюнок від Dylus et al. 1 Опубліковано під ліцензією CC BY 4.0. Малюнок був обрізаний з оригіналу. |
Загальна картина: Долі клітин поступово обмежуються в міру розвитку
В ході розвитку повинні бути уточнені всі органи, тканини, клітини-типи. У тварин це досягається повторним обмеженням долі 2. Наприклад, тотипотентний ембріон проходить кілька раундів ділення з утворенням бластули (кульки клітин). Деякі з цих клітин візьмуть на себе мезодермальну долю. У цей момент вони можуть стати будь-яким типом мезодермальних клітин. У деяких тварин експерименти були проведені, щоб показати, що родова мезодермальна клітина може бути індукована сусідами розвиватися в величезну різноманітність типів мезодермальних клітин 3. Однак з часом ці мезодермальні клітини діляться, зазнають змін форми, мігрують та диференціюються. Коли вони отримують сигнали зі свого середовища (переважно клітин навколо них), вони диференціюються на більш конкретні типи клітин. Наприклад, на малюнку 3 показана мезодермальна клітина, що диференціюється на м'язові, кісткові та кров'яні клітини. З часом різні м'язові попередники (міогенні) клітини будуть диференціюватися на різні типи м'язів, наприклад, гладкі та поперечно-смугасті. Кожне з цих рішень про клітинну долю залежить від схеми експресії гена в клітині, яка залежить від історії клітини та її сусідів.

Ми називаємо цей тип прогресивної диференціації «ієрархічним» з більш загальними типами клітин (наприклад, мезодерма, рис. 3) у верхній частині ієрархії та більш конкретними типами клітин (як нейтрофіли) внизу 4. У міру розвитку ми рухаємося вниз по ієрархії. Розробка починається специфікація осі, ці осі незабаром зчитуються в регіони - наприклад, голова, тулу/черевце і хвіст. У межах цих різних областей тканини уточнюються спочатку на рівні зародкових шарів, а пізніше на більш конкретних рівнях. Наприклад, ектодерма розщеплюється на нейроектодерму і епідермальну ектодерму в залежності від розташування тканини. Далі уточнюються клітини-типи всередині тканин. Наприклад, нейроектодерма може стати нейронами або гліальними клітинами. Обидва ці типи клітин пізніше диференціюються на багато специфічних типів нейронів і глії. У цей момент клітини більш-менш невиліковно диференційовані - уточнюються їх дорослі долі, і вони не можуть повернутися до плюрипотентних форм. Однак більшість клітин все ще сприйнятливі до навколишнього середовища і можуть реагувати на сигнали навколишнього середовища навіть після того, як вони невиліковно диференційовані. Ми називаємо це клітинною пластичністю.
GRN на малюнку 2 відображає цю ієрархію, причому ранні діючі гени вгорі визначають більш загальні тканини, а пізніше діючі гени внизу визначають конкретні типи клітин. Однак важливо зазначити, що гени, що діють на цих різних ієрархічних рівнях, повторно використовуються на інших рівнях ієрархії та в інших тканині, що розвиваються. Насправді основні частини GRN та шляхи передачі сигналів повторно використовуються протягом усього розвитку та мають різні виходи залежно від їх контексту.