23.1: Еукаріотичне походження


- Last updated
- Oct 23, 2022
- Save as PDF

Навички для розвитку
- Перерахуйте об'єднуючі характеристики еукаріотів
- Опишіть, що знають вчені про походження еукаріотів на основі останнього спільного предка
- Поясніть ендосимбіотичну теорію
Живі істоти діляться на три великі групи: Археї, Бактерії та Еукарія. Перші дві мають прокаріотичні клітини, а треті містять всі еукаріоти. Доступний відносно розріджений запис викопних копалин, щоб допомогти розрізнити, як виглядали перші члени кожної з цих родовищ, тому цілком можливо, що всі події, які призвели до останнього спільного предка існуючих еукаріотів, залишаться невідомими. Однак порівняльна біологія існуючих організмів та обмежений запис викопних копалин дають деяке уявлення про історію Еукарії.
Найраніші знайдені скам'янілості виявляються бактеріями, швидше за все ціанобактеріями. Їм близько 3,5 мільярда років і впізнавані через їх відносно складну структуру і, для прокаріотів, відносно великих клітин. Більшість інших прокаріотів мають невеликі клітини розміром 1 або 2 мкм, і їх важко буде вибрати як скам'янілості. Більшість живих еукаріотів мають клітини розміром 10 мкм або більше. Структури такого розміру, які можуть бути скам'янілостями, з'являються в геологічному рекорді близько 2,1 мільярда років тому.
характеристика еукаріотів
Дані цих скам'янілостей привели порівняльних біологів до висновку, що живі еукаріоти - все це нащадки єдиного спільного предка. Картування характеристик, знайдених у всіх основних групах еукаріотів, виявляє, що наступні характеристики повинні були бути присутніми в останньому спільному предку, оскільки ці характеристики присутні принаймні у деяких членів кожної основної лінії.
- Клітини з ядрами, оточені ядерною оболонкою з ядерними порами. Це єдина характеристика, яка є і необхідною, і достатньою для визначення організму як еукаріот. Всі збереглися еукаріоти мають клітини з ядрами.
- Мітохондрії. Деякі збереглися еукаріоти мають дуже зменшені залишки мітохондрій у своїх клітині, тоді як інші члени їх родовищ мають «типові» мітохондрії.
- Цитоскелет, що містить структурні та рухливі компоненти, звані актиновими мікрофіламентами та мікротрубочками. Всі існуючі еукаріоти мають ці цитоскелетні елементи.
- Джгутики і вії, органели, пов'язані з рухливістю клітин. Деякі збереглися еукаріот позбавлені джгутиків і/або вій, але вони походять від предків, які володіли ними.
- Хромосоми, кожна з яких складається з лінійної молекули ДНК, згорнутої навколо основних (лужних) білків, званих гістонами. Нечисленні еукаріоти з хромосомами, які не мають гістонів, явно еволюціонували від предків, які їх мали.
- Мітоз, процес ядерного поділу, при якому репліковані хромосоми поділяються і відокремлюються за допомогою елементів цитоскелета. Мітоз повсюдно присутній у еукаріот.
- Секс, процес генетичної рекомбінації, унікальний для еукаріотів, при якому диплоїдні ядра на одному етапі життєвого циклу піддаються мейозу з отриманням гаплоїдних ядер та подальшої каріогамії, стадії, коли два гаплоїдні ядра зливаються разом, створюючи диплоїдне ядро зиготи.
- Члени всіх основних родовищ мають клітинні стінки, і може бути розумним зробити висновок, що останній загальний предок може зробити клітинні стінки на певному етапі свого життєвого циклу. Однак недостатньо відомо про клітинні стінки еукаріотів та їх розвиток, щоб знати, скільки гомології існує серед них. Якщо останній загальний предок міг зробити клітинні стінки, то зрозуміло, що ця здатність повинна була бути втрачена в багатьох групах.
Ендосимбіоз і еволюція еукаріотів
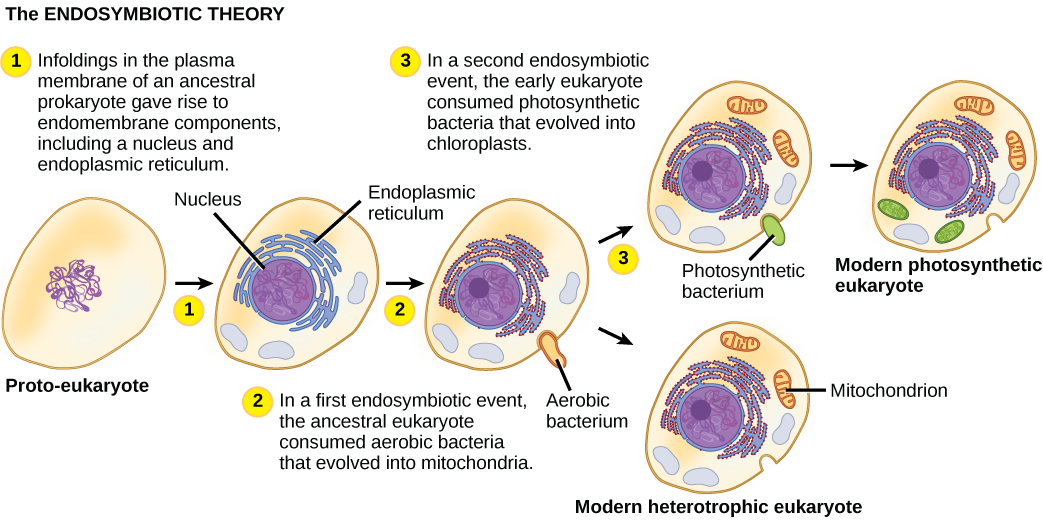
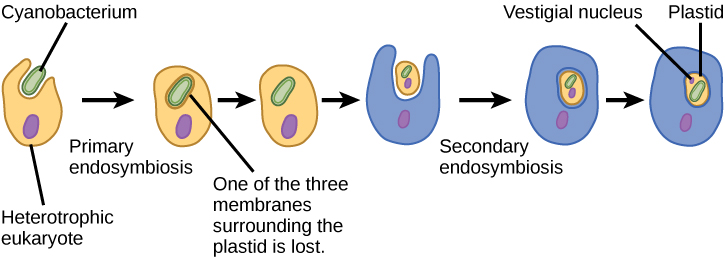
Для того щоб повністю зрозуміти еукаріотичні організми, необхідно розуміти, що всі існуючі еукаріоти є нащадками химерного організму, який був складом клітини-господаря і клітини (ів) альфа-протеобактерії, яка «проживала» всередині неї. Ця головна тема походження еукаріотів відома як ендосимбіоз, одна клітина охоплює іншу таку, що охоплена клітина виживає, і обидві клітини отримують користь. Протягом багатьох поколінь симбіотичні відносини можуть призвести до двох організмів, які залежать один від одного настільки повністю, що жоден не міг вижити самостійно. Ендосимбіотичні події, ймовірно, сприяли виникненню останнього спільного предка сучасних еукаріотів і пізнішої диверсифікації в певних родовищах еукаріотів (рис.23.1.4). Перш ніж пояснити це далі, необхідно розглянути метаболізм у прокаріотів.
Прокаріотичний метаболізм
Багато важливих обмінних процесів виникли у прокаріотів, а деякі з них, такі як фіксація азоту, ніколи не зустрічаються у еукаріотів. Процес аеробного дихання зустрічається у всіх основних лініях еукаріот, і локалізується він в мітохондріях. Аеробне дихання зустрічається і в багатьох родовищах прокаріотів, але воно присутнє не у всіх з них, і багато форм доказів говорять про те, що такі анаеробні прокаріоти ніколи не здійснювали аеробного дихання, як і їхні предки.
Хоча сьогоднішня атмосфера становить близько однієї п'ятої молекулярного кисню (O 2), геологічні дані показують, що спочатку їй не вистачало O 2. Без кисню аеробного дихання не очікувалося б, а живі істоти покладалися б на бродіння замість цього. У якийсь момент раніше, приблизно 3,5 мільярда років тому, деякі прокаріоти почали використовувати енергію від сонячного світла для живлення анаболічних процесів, які зменшують вуглекислий газ з утворенням органічних сполук. Тобто у них розвивалася здатність до фотосинтезу. Водень, отриманий з різних джерел, був захоплений за допомогою світлових реакцій для зменшення фіксованого вуглекислого газу в циклі Кальвіна. Група грамнегативних бактерій, що породили ціанобактерії, використовували воду як джерело водню і виділяли O 2 як продукт життєдіяльності.
Зрештою, кількість фотосинтетичного кисню накопичується в деяких середовищах до рівнів, що представляють ризик для живих організмів, оскільки це може пошкодити багато органічних сполук. Еволюціонували різні обмінні процеси, які захищали організми від кисню, один з яких, аеробне дихання, також генерував високий рівень АТФ. Він став широко присутнім серед прокаріотів, в тому числі і в групі, яку ми зараз називаємо альфа-протеобактеріями. Організми, які не набули аеробного дихання, повинні були залишатися в безкисневих середовищах. Спочатку багаті киснем середовища були, ймовірно, локалізовані навколо місць, де діяли ціанобактерії, але приблизно 2 мільярди років тому геологічні дані показують, що кисень нарощувався до більш високих концентрацій в атмосфері. Рівень кисню, подібний до сьогоднішніх рівнів, виник лише протягом останніх 700 мільйонів років.
Нагадаємо, що перші скам'янілості, які, на нашу думку, є еукаріотами, становлять близько 2 мільярдів років, тому вони з'явилися, коли рівень кисню збільшувався. Також нагадаємо, що всі дійшли до наших днів еукаріоти походять від предка з мітохондріями. Ці органели вперше спостерігали легкі мікроскопісти наприкінці 1800-х років, де вони виявилися дещо черв'ячними структурами, які, здавалося, рухалися в клітці. Деякі ранні спостерігачі припускали, що це можуть бути бактерії, що живуть всередині клітин господаря, але ці гіпотези залишилися невідомими або відхиленими в більшості наукових спільнот.
Ендосимбіотична теорія
Оскільки клітинна біологія розвивалася в ХХ столітті, стало зрозуміло, що мітохондрії - це органели, відповідальні за вироблення АТФ за допомогою аеробного дихання. У 1960-х роках американський біолог Лінн Маргуліс розробила ендосимбіотичну теорію, яка стверджує, що еукаріоти, можливо, були продуктом однієї клітини, що охоплює іншу, одна живе в іншій і еволюціонує з часом, поки окремі клітини більше не були впізнавані як такі. У 1967 році Маргуліс представила нову роботу з теорії і обґрунтувала свої висновки за допомогою мікробіологічних доказів. Хоча робота Маргуліса спочатку зустрічалася з опором, ця колись революційна гіпотеза зараз широко (але не повністю) прийнята, при цьому робота просувається над розкриттям кроків, залучених до цього еволюційного процесу, та ключових гравців, залучених до цього. Ще багато ще належить відкрити про походження клітин, які зараз складають клітини у всіх живих еукаріотів.
Загалом, стало зрозуміло, що багато наших ядерних генів та молекулярних механізмів, відповідальних за реплікацію та експресію, виявляються тісно пов'язаними з тими, що знаходяться в Археї. З іншого боку, метаболічні органели та гени, відповідальні за багато процесів збору енергії, мали своє походження з бактерій. Багато що ще належить з'ясувати про те, як відбулися ці відносини; це продовжує залишатися захоплюючим полем відкриття в біології. Наприклад, невідомо, чи відбулася ендосимбіотична подія, яка призвела до мітохондрій до чи після того, як клітина-господар мала ядро. Такі організми були б серед вимерлих попередників останнього спільного предка еукаріотів.
Мітохондрії
Однією з головних ознак, що відрізняють прокаріотів від еукаріотів, є наявність мітохондрій. Еукаріотичні клітини можуть містити від однієї до декількох тисяч мітохондрій, залежно від рівня споживання енергії клітиною. Кожен мітохондріон вимірює довжину від 1 до 10 або більше мікрометрів і існує в клітці як органела, яка може бути яйцеподібною до черв'ячної форми або хитромудро розгалуженою (рис.23.1.1). Мітохондрії виникають внаслідок поділу існуючих мітохондрій; вони можуть зливатися разом; і вони можуть переміщатися всередині клітини шляхом взаємодії з цитоскелетом. Однак мітохондрії не можуть вижити поза клітиною. Оскільки атмосфера насичувалася киснем шляхом фотосинтезу, і в міру розвитку успішних аеробних прокаріотів, дані свідчать про те, що родова клітина з деякою мембранною компартменталізацією охопила вільноживучий аеробний прокаріот, зокрема альфа-протеобактерію, тим самим надаючи клітині-господареві можливість використовувати кисень для вивільнення енергії, що зберігається в поживних речовині. Альфа-протеобактерії - це велика група бактерій, яка включає в себе види, симбіотичні з рослинами, хвороботворні організми, які можуть заразити людину за допомогою кліщів, і багато вільно живуть види, які використовують світло для отримання енергії. Кілька ліній доказів підтверджують, що мітохондрії походять від цієї ендосимбіотичної події. Більшість мітохондрій мають форму альфа-протеобактерій і оточені двома мембранами, що призвело б до того, коли один пов'язаний з мембраною організм був поглинаний у вакуоль іншим організмом, пов'язаним з мембраною. Внутрішня мембрана мітохондрій обширна і включає значні вкладення, звані кристами, які нагадують текстуровану зовнішню поверхню альфа-протеобактерій. Матрикс і внутрішня мембрана багаті ферментами, необхідними для аеробного дихання.
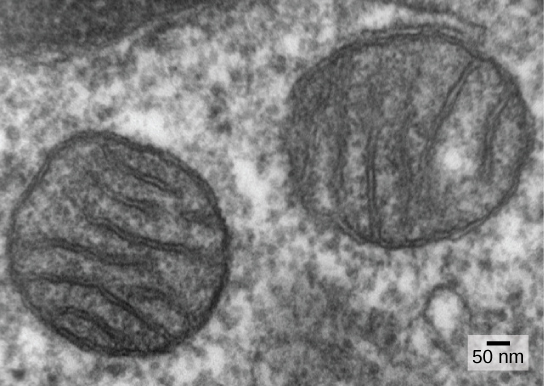
Мітохондрії діляться самостійно процесом, який нагадує бінарне ділення у прокаріотів. Зокрема, мітохондрії не утворюються з нуля (de novo) еукаріотичною клітиною; вони розмножуються всередині неї і розподіляються з цитоплазмою, коли клітина ділиться або зливається дві клітини. Тому, хоча ці органели високо інтегровані в еукаріотичну клітину, вони все ще розмножуються так, ніби є незалежними організмами всередині клітини. Однак їх розмноження синхронізується з активністю і поділом клітини. Мітохондрії мають власну (зазвичай) кругову ДНК-хромосому, яка стабілізується прикріпленнями до внутрішньої мембрани і несе гени, схожі на гени, експресовані альфа-протеобактеріями. Мітохондрії також мають особливі рибосоми і переносять РНК, які нагадують ці компоненти у прокаріотів. Всі ці особливості підтримують те, що мітохондрії колись були вільноживучими прокаріотами.
Мітохондрії, які здійснюють аеробне дихання, мають власні геноми, з генами, подібними до генів у альфа-протеобактерій. Однак багато генів для дихальних білків розташовані в ядрі. Коли ці гени порівнюються з генами інших організмів, вони, здається, мають альфа-протеобактеріальне походження. Крім того, в деяких еукаріотичних групах такі гени знаходяться в мітохондріях, тоді як в інших групах вони знаходяться в ядрі. Це було інтерпретовано як доказ того, що гени були перенесені з ендосимбіонтної хромосоми в геном господаря. Ця втрата генів ендосимбіонтом, ймовірно, є одним з пояснень, чому мітохондрії не можуть жити без господаря.
Деякі живі еукаріоти анаеробні і не можуть вижити в присутності занадто великої кількості кисню. Деяким, здається, не вистачає органел, які можна було б визнати мітохондріями. У 1970-х до початку 1990-х років багато біологів припустили, що деякі з цих еукаріотів походять від предків, чиї лінії відрізнялися від лінії мітохондріонсодержащих еукаріотів до того, як стався ендосимбіоз. Однак пізніші висновки свідчать про те, що знижені органели зустрічаються у більшості, якщо не у всіх, анаеробних еукаріотів, і що всі еукаріоти, здається, несуть у своїх ядрах деякі гени, які мають мітохондріальне походження. Крім аеробної генерації АТФ, мітохондрії мають ще кілька метаболічних функцій. Однією з цих функцій є генерування скупчень заліза та сірки, які є важливими кофакторами багатьох ферментів. Такі функції часто пов'язані з редукованими мітохондріями органелами анаеробних еукаріотів. Тому більшість біологів сприймають, що останній загальний предок еукаріот мав мітохондрії.
Пластиди
Деякі групи еукаріотів є фотосинтетичними. Їх клітини містять, крім стандартних еукаріотичних органел, ще один вид органел під назвою пластида. Коли такі клітини проводять фотосинтез, їх пластиди багаті пігментом хлорофілом а і рядом інших пігментів, званих допоміжними пігментами, які беруть участь у збирання енергії зі світла. Фотосинтетичні пластиди називаються хлоропластами (рис.23.1.2).
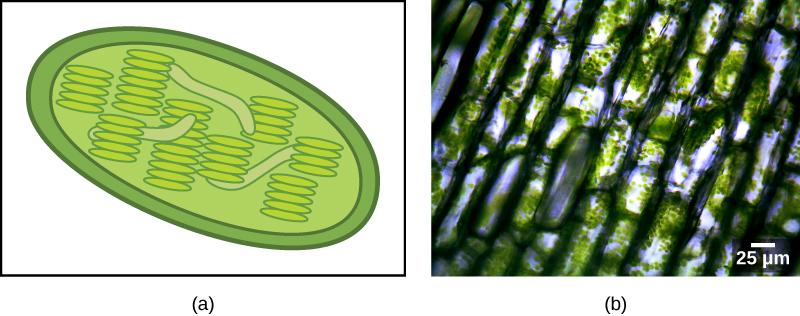
Як і мітохондрії, пластиди, здається, мають ендосимбіотичне походження. Цю гіпотезу також відстоювала Лінн Маргуліс. Пластиди походять з ціанобактерій, які мешкали всередині клітин предкового, аеробного, гетеротрофного еукаріота. Це називається первинним ендосимбіозом, а пластиди первинного походження оточені двома мембранами. Найкращим свідченням є те, що це траплялося двічі в історії еукаріотів. В одному випадку загальний предок основної родості/супергрупи Archeaplastida взяв на себе ціанобактеріальний ендосимбіонт; в іншому родоначальник малого амебоїдного ризарійського таксона, Paulinella, взяв на себе інший ціанобактеріальний ендосимбіонт. Майже всі фотосинтезуючі еукаріоти походять від першої події, і лише пара видів походить від іншого.
Ціанобактерії - це група грамнегативних бактерій з усіма умовними структурами групи. Однак, на відміну від більшості прокаріотів, вони мають великі, внутрішні мембранно-зв'язані мішечки, звані тилакоїдами. Хлорофіл є компонентом цих мембран, як і багато білків світлових реакцій фотосинтезу. Ціанобактерії також мають стінку пептидоглікану і ліпополісахаридний шар, пов'язаний з грамнегативними бактеріями.
Хлоропласти первинного походження мають тилакоїди, кругову хромосому ДНК та рибосоми, подібні до ціанобактерій. Кожен хлоропласт оточений двома мембранами. У групі Археопластіда під назвою глаукофіти, а в Paulinella тонкий пептидоглікановий шар присутній між зовнішньою і внутрішньою пластидними мембранами. Всі інші пластиди позбавлені цієї реліктової ціанобактеріальної стінки. Вважається, що зовнішня мембрана, що оточує пластид, походить від вакуолі в господаря, а внутрішня мембрана, як вважається, походить від плазматичної мембрани симбіонта.
Є також, як і у випадку з мітохондріями, вагомі докази того, що багато генів ендосимбіонта були перенесені в ядро. Пластиди, як і мітохондрії, не можуть жити самостійно поза господарем. Крім того, як і мітохондрії, пластиди походять від поділу інших пластид і ніколи не будуються з нуля. Дослідники припустили, що ендосимбіотична подія, яка призвела до Архепластиди, сталася від 1 до 1,5 мільярда років тому, принаймні через 5 сотень мільйонів років після того, як викопний запис свідчить про наявність еукаріотів.
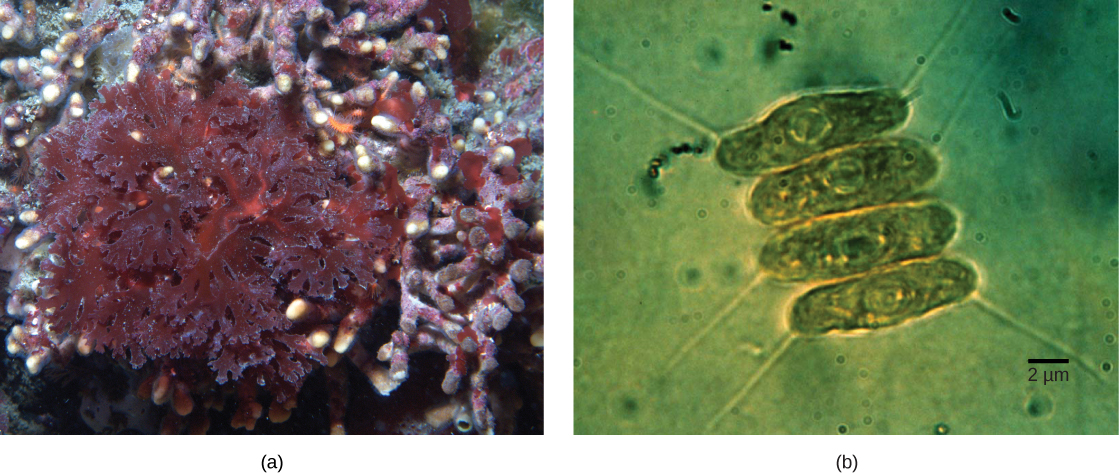
Не всі пластиди в еукаріотів отримані безпосередньо з первинного ендосимбіозу. Деякі з основних груп водоростей стали фотосинтетичними шляхом вторинного ендосимбіозу, тобто шляхом прийняття або зелених водоростей, або червоних водоростей (обидва від Archaepastida) як ендосимбіонти (рис.23.1.3). Численні мікроскопічні та генетичні дослідження підтвердили цей висновок. Вторинні пластиди оточені трьома і більше мембранами, а деякі вторинні пластиди навіть мають чіткі залишки ядра ендосимбіотичних водоростей. Інші не «зберегли» ніяких залишків. Бувають випадки, коли ендосимбіотичні події третинного або вищого порядку є найкращим поясненням пластид у деяких еукаріотів.
Мистецтво З'єднання
Які докази того, що мітохондрії були включені в родову еукаріотичну клітину перед хлоропластами?
Еволюційний зв'язок: вторинний ендосимбіоз у хлорахніофітів
Ендосимбіоз включає в себе одну клітину, що охоплює іншу, щоб з часом виробляти співрозвинені відносини, в яких жодна клітина не могла вижити поодинці. Наприклад, хлоропласти червоних і зелених водоростей походять від поглинання фотосинтетичної ціанобактерії раннім прокаріотом.
Це призводить до питання про можливість того, щоб клітина, що містить ендосимбіонт, до себе стати поглинутою, в результаті чого виникає вторинний ендосимбіоз. Молекулярні та морфологічні дані свідчать про те, що протести хлорахніофіту походять від вторинної ендосимбіотичної події. Хлорарахніофіти - рідкісні водорості, корінні з тропічних морів і піску, які можна класифікувати на ризарійську супергрупу. Хлорахіофіти подовжують тонкі цитоплазматичні нитки, з'єднуючись між собою з іншими хлорахніофітами, в цитоплазматичну мережу. Вважається, що ці протести виникли, коли еукаріот охопив зелену водорость, остання з яких вже встановила ендосимбіотичний зв'язок з фотосинтетичною ціанобактерією (рис.23.1.5).
Кілька ліній доказів підтверджують, що хлорахіофіти еволюціонували з вторинного ендосимбіозу. Хлоропласти, що містяться в ендосимбіонтах зелених водоростей, все ще здатні до фотосинтезу, роблячи хлорахіофіти фотосинтетичними. Ендосимбіонт із зелених водоростей також демонструє затримане вестигіальне ядро. Насправді, виявляється, що хлорахніофіти є продуктами еволюційно недавнього вторинного ендосимбіотичного події. Пластіди хлорахінофітів оточені чотирма мембранами: перші дві відповідають внутрішній і зовнішній мембранах фотосинтетичної ціанобактерії, третя відповідає зеленої водорості, а четверта відповідає вакуолі, яка оточувала зелені водорості, коли вона була охоплена предок хлорахніофіту. В інших лініях, які брали участь у вторинному ендосимбіозі, навколо пластид можна ідентифікувати лише три мембрани. В даний час це випрямляється як послідовна втрата мембрани під час еволюції.
Процес вторинного ендосимбіозу не є унікальним для хлорахніофітів. Насправді вторинний ендосимбіоз зелених водоростей також призвів до евгленідних протестів, тоді як вторинний ендосимбіоз червоних водоростей призвів до еволюції динофлагеллатів, апікомплексів і страменопілів.
Резюме
Найдавніше викопне свідоцтво еукаріотів становить близько 2 мільярдів років. Скам'янілості старше цього все здаються прокаріотами. Цілком ймовірно, що сьогоднішні еукаріоти походять від предка, який мав прокаріотичну організацію. Останній загальний предок сьогоднішньої Еукарії мав кілька характеристик, включаючи клітини з ядрами, які розділилися мітотично і містили лінійні хромосоми, де ДНК була пов'язана з гістонами, цитоскелетом і ендомембранною системою, а також здатність робити вії/джгутики протягом принаймні частини свого життя цикл. Це було аеробним, оскільки в ньому були мітохондрії, які були результатом аеробної альфа-протеобактерії, яка жила всередині клітини-господаря. Чи мав у цього господаря ядро на момент початкового симбіозу, залишається невідомим. Останній загальний предок, можливо, мав клітинну стінку протягом принаймні частини свого життєвого циклу, але для підтвердження цієї гіпотези потрібні додаткові дані. Сьогоднішні еукаріоти дуже різноманітні за своїми формами, організацією, життєвими циклами та кількістю клітин на людину.
Мистецькі зв'язки
Малюнок23.1.4: Які докази того, що мітохондрії були включені в родову еукаріотичну клітину перед хлоропластами?
- Відповідь
-
Всі еукаріотичні клітини мають мітохондрії, але не всі еукаріотичні клітини мають хлоропласти.
Глосарій
- ендосимбіоз
- поглинання однієї клітини всередині іншої таким чином, що охоплена клітина виживає, і обидві клітини виграють; процес, відповідальний за еволюцію мітохондрій і хлоропластів в еукаріотів
- ендосимбіотична теорія
- теорія, яка стверджує, що еукаріоти, можливо, були продуктом однієї клітини, що охоплює іншу, одну, що живе в іншій, і розвивається з часом, поки окремі клітини більше не були впізнавані як такі
- пластид
- одна з групи споріднених органел у клітині рослин, які беруть участь у зберіганні крохмалю, жирів, білків та пігментів