9.3: Регуляція експресії генів у еукаріотів
- Page ID
- 5666

Останні оцінки полягають у тому, що людська клітина, еукаріотична клітина, містить близько 21 000 генів. Деякі з них весь час виражаються у всіх клітинок. Ці так звані гени домашнього господарства відповідають за рутинні метаболічні функції (наприклад, дихання), загальні для всіх клітин. Деякі виражаються як клітина входить в певний шлях диференціації. Деякі виражаються весь час тільки в тих осередках, які диференціювалися певним чином. Наприклад, плазматична клітина безперервно експресує гени для синтезованого нею антитіла. Деякі виражаються лише як зміни умов навколо і в клітині. Наприклад, прихід гормону може включити (або вимкнути) певні гени в цій клітині.
Як регулюється експресія генів? Існує кілька методів, які використовуються еукаріотами.
- Зміна швидкості транскрипції гена. Це найважливіша і широко використовувана стратегія.
- Однак еукаріоти доповнюють транскрипційну регуляцію кількома іншими методами:
- Зміна швидкості обробки стенограми РНК, перебуваючи в ядрі.
- Зміна стабільності молекул месенджерної РНК (мРНК); тобто швидкість їх деградації.
- Зміна ефективності, з якою рибосоми переводять мРНК в поліпептид.
Гени, що кодують білок, мають
- екзони, послідовність яких кодує поліпептид
- інтрони, які будуть видалені з мРНК перед її перекладом
- сайт початку транскрипції
- промоутери
- базальний або основний промоутер, розташований в межах приблизно 40 пар бази (bp) від стартового сайту
- Промоутери «вгору за течією», які можуть поширюватися на цілих 200 б.п. далі вгору за течією
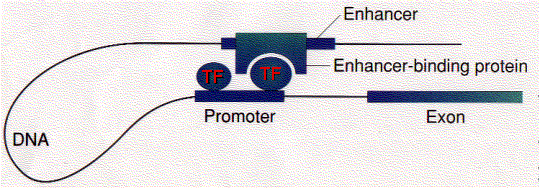
- підсилювачі
- глушники
Сусідні гени часто розділені ізолятором, який допомагає їм уникнути перехресних розмов між промоторами та підсилювачами один одного (та/або глушниками).
Сайт початку транскрипції
Тут зв'язується молекула РНК-полімерази II (пол II, також відома як RNAP II). Пол II являє собою комплекс з 12 різних білків (показаний на малюнку жовтим кольором з накладеними на нього невеликими кольоровими колами). Стартова ділянка - це місце, де починається транскрипція гена в РНК.
Основний промоутер
Всі еукаріотичні гени містять промотор ядра. Одним із поширених прикладів є послідовність основ (наприклад, TATAAAAAA), яка називається коробкою TATA. Він пов'язаний великим комплексом з якихось 50 різних білків, в тому числі
- Фактор транскрипції IID (TFIID), який є комплексом
- Тата-зв'язуючий білок (ТБП), який розпізнає і зв'язується з коробкою TATA
- 13 інших білкових факторів, які зв'язуються з ТБП, один з одним і (деякі з них) з ДНК.
- Фактор транскрипції IIB (TFIIB), який пов'язує як ДНК, так і пол II.
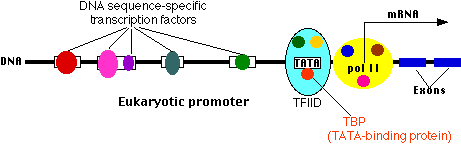
Багато різних генів і багато різних типів клітин мають однакові фактори транскрипції - не тільки ті, що зв'язуються з основним промотором, але навіть деякі з тих, що зв'язуються вище за течією. Те, що перетворюється на певний ген у певній клітині, ймовірно, унікальне поєднання промоторних сайтів та факторів транскрипції, які вибираються.
Аналогія
Ряди замкових коробок в банку дають корисну аналогію. Для відкриття якогось конкретного ящика в кімнаті потрібно два ключа:
- Ваш ключ, шаблон виїмок якого відповідає лише замку призначеної вам коробки (= промоутер вище за течією), але який не може розблокувати коробку без
- Ключ, який несе співробітник банку, який може активувати механізм розблокування будь-якого ящика (= основний промоутер), але сам по собі не може відкрити жодної скриньки.
Ефект гормонів
Ці петлі стабілізуються за допомогою білка, позначеного CTCF («фактор зв'язування CCCTC»; названий на честь нуклеотидної послідовності, з якою він зв'язується). CTCF на одній ділянці на ДНК утворює димер з CTCF на іншому місці на ДНК, що зв'язує дві області разом. CTCF має 11 цинкових пальців. Вони також можуть бути стабілізовані когезином - тим самим білковим комплексом, який утримує сестринські хроматиди разом під час мітозу та мейозу.
Майкл Ботчан і його колеги підготували наочні докази цієї моделі енхансерного дії. Вони створили штучну молекулу ДНК з
- кілька (4) промоутерських сайтів для Sp1 близько 300 баз з одного кінця. Sp1 - це фактор транскрипції цинкового пальця, який зв'язується з послідовністю 5' GGGCGG 3', знайденої в промоторах багатьох генів, особливо «господарських» генів.
- кілька (5) підсилювачів сайтів близько 800 баз з іншого кінця. Вони пов'язані підсилювачем зв'язуючого білка, позначеного Е2.
- 1860 пар основ ДНК між ними.

Значення «зациклювання»
Циклічне хромосом, яке наближає підсилювачі до промоторів (і промоторів, близьких до інших промоторів), здається, є механізмом забезпечення експресії (або гальмування) груп генів, які повинні виконувати разом. Відповідь клітини на надходження сигналу (наприклад, гормону) може включати включення (або виключення) сотень різних генів, продукти яких повинні вироблятися скоординовано, щоб клітина відповідала належним чином. Динамічний рух частин хромосоми, що несуть відповідні генні локуси в «фабрику транскрипції», може бути механізмом для досягнення цього. Якщо так, ми бачимо еукаріотичний еквівалент скоординованої експресії генів, що забезпечується оперонами у бактерій.
Глушники
Глушники - це контрольні ділянки ДНК, які, як і підсилювачі, можуть розташовуватися на тисячах пар основ від гена, який вони контролюють. Однак, коли транскрипційні фактори зв'язуються з ними, експресія контрольованого ними гена пригнічується.
Ізолятори
Як ви можете бачити вище, підсилювачі можуть включати промотори генів, розташованих за тисячами пар основ. Ізолятори перешкоджають енхансеру від неналежного зв'язування та активації промотора якогось іншого гена в тій же області хромосоми..
Ізолятори бувають
- розтяжки ДНК (всього 42 пари основ можуть зробити трюк)
- розташований між
- підсилювач (и) і промоутер (и) або
- глушник (и) і промоутер (и)

Енхансер для промотора гена дельта-ланцюга гамма/дельта-Т-клітинного рецептора антигену (TCR) розташований близько до промотора альфа-ланцюга альфа/бета TCR (на хромосомі 14 у людини). Т-клітина повинна вибирати між тим чи іншим. Між промотором альфа-генів та промотором дельта-гена є ізолятор, який гарантує, що активація одного не поширюється на інший.
Всі ізолятори, виявлені до цих пір у хребетних, працюють тільки при зв'язці білком CTCF. Інший приклад: У ссавців (мишей, людей, свиней) активний лише алель для інсуліноподібного фактора росту-2 (IGF2), успадкований від батька; що успадковано від матері не є — явище під назвою імпринтінг.
Механізм: алель матері має ізолятор між промотором IGF2 і підсилювачем. Так само і алель батька, але в його випадку ізолятор був метильований. CTCF більше не може зв'язуватися з ізолятором, і тому підсилювач тепер може вільно включати промоутер IGF2 батька.
Багато з комерційно важливих сортів свиней були виведені для того, щоб містити ген, який збільшує співвідношення скелетних м'язів до жиру. Цей ген був секвенований і виявляється алелем IGF2, який містить єдину точкову мутацію в одному з його інтронів. Свині з цією мутацією виробляють більш високий рівень IGF2 мРНК в їх скелетних м'язах (але не в їх печінці). Це говорить нам про те, що:
- Мутації не повинні бути в білковій кодуючій частині гена, щоб вплинути на фенотип.
- Мутації в некодуючих частинок гена можуть впливати на те, як цей ген регулюється (тут зміна м'язів, але не в печінці).