5.1: Трансформація бактерій
- Page ID
- 5764

Бактерії не мають статевого розмноження в тому сенсі, що це роблять еукаріоти. У них немає чергування диплоїдних і гаплоїдних поколінь, ніяких гамет і мейозу. Однак суть сексу полягає в генетичній рекомбінації, і бактерії мають три механізми для цього: перетворення, кон'югація та трансдукція.
Трансформація
Багато бактерій можуть набувати нових генів, приймаючи молекули ДНК (наприклад, плазміду) зі свого оточення. Здатність навмисно трансформувати бактерію кишкової палички зробила можливим клонування багатьох генів, включаючи гени людини, і розвиток біотехнологічної галузі. Перша демонстрація бактеріальної трансформації була зроблена за допомогою Streptococcus pneumoniae і призвела до відкриття, що ДНК є речовиною генів. Шляхи, що ведуть до цього епохального відкриття, розпочався в 1928 році з роботи англійського бактеріолога Фреда Гріффіта.


Клітини S. pneumoniae (також відомі як пневмокок) зазвичай оточені клейкою капсулою, виготовленою з полісахариду. При вирощуванні на поверхні твердого культурального середовища капсула змушує колонії мати блискучий, гладкий вигляд. Ці клітини називаються «S» клітинами. Однак після тривалого культивування на штучному середовищі деякі клітини втрачають здатність утворювати капсулу, а поверхня їх колоній зморшкувата і шорстка («R»). При втраті своєї капсули бактерії також втрачають свою вірулентність. Ін'єкція одного S пневмокока в мишу вб'є миша через 24 години або близько того. Але ін'єкція понад 100 мільйонів (100 х 10 6) R клітин абсолютно нешкідлива.

Причина? Капсула запобігає поглинанню та знищенню пневмококів шляхом очищення клітин, нейтрофілів та макрофагів в організмі. Форми R повністю перебувають у владі фагоцитів. Пневмококи також зустрічаються понад 90 різних типів: I, II, III і так далі. Види відрізняються хімією їх полісахаридної капсули. На відміну від випадкового зсуву S -> R, тип організму постійний. Миші, які вводять кілька S клітин, скажімо, пневмококів II типу, незабаром їх тіла кишать нащадковими клітинами того ж типу.
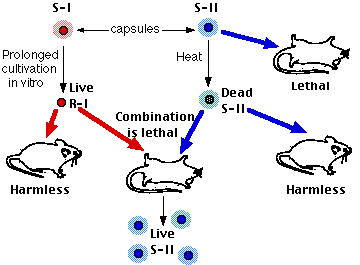
Однак Гріффіт виявив, що коли живі R-клітини (які повинні були бути нешкідливими) та мертві S-клітини (які також повинні були бути нешкідливими) вводяться разом, миша захворіла і живі S-клітини могли бути відновлені з її тіла. Крім того, тип клітин, витягнутих з тіла миші, визначався типом мертвих S-клітин. В експерименті, показаному вище, ін'єкції живих R-I клітин і мертвих S-II клітин виробляли вмираючу мишу з її тілом, наповненим живими S-II пневмококами. Клітини S-II залишилися вірними своєму новому типу. Щось у мертвих S-II клітині зробило постійну зміну фенотипу R-I клітин. Процес отримав назву трансформації.
Освальд Ейвері та його колеги з Інституту Рокфеллера в Нью-Йорку врешті-решт показали, що «щось» - це ДНК. Переслідуючи відкриття Гріффіта, вони виявили, що вони можуть принести приблизно такий же вид трансформації in vitro за допомогою екстракту бактеріальних клітин. Лікували цим екстрактом:
- фермент для руйнування полісахариду капсули (S-III в своїх дослідах)
- розчинники для видалення всіх ліпідів
- трипсин і хімотрипсин для знищення будь-яких залишкових білків
- РНАза для знищення РНК
Вони виявили, що це не руйнує здатність їх екстрактів перетворювати бактерії. Однак лікування екстрактів ДНК для знищення ДНК в них скасувало їх трансформуючу активність. Таким чином, ДНК була єдиним матеріалом у мертвих клітині, здатним перетворювати клітини від одного типу до іншого. ДНК була речовиною генів.
Хоча хімічний склад капсули визначається генами, взаємозв'язок непряма. ДНК транскрибується в РНК, а РНК перетворюється на білки. Фенотип пневмококів — хімічний склад полісахаридної капсули — визначається особливими ферментами (білками), які використовуються в синтезі полісахаридів. Ейвері та його колеги Колін Маклауд і Маклін Маккарті опублікували свої епохальні знахідки 1 лютого 1944 року. На жаль, важливість їх відкриття не була достатньо оцінена вченими в цілому і Нобелівським комітетом зокрема, і Ейвері помер до того, як їх роботи змогли удостоїтися Нобелівської премії. (Нобелівські премії ніколи не вручаються посмертно.)
відмінювання
Деякі бактерії, наприклад, кишкова паличка, можуть передавати частину своєї хромосоми реципієнту, з яким вони знаходяться в безпосередньому контакті. У міру того, як донор реплікує свою хромосому, копія вводиться реципієнту. У будь-який момент, коли донор і реципієнт розділяються, передача генів припиняється. Ті гени, які успішно здійснили поїздку, замінюють їх еквіваленти в хромосомі реципієнта.
Кон'югація може відбуватися лише між клітинами протилежних типів спаровування: донор (або «самець») несе фактор фертильності (F +), а реципієнт («самка») - ні (F −). Фактор фертильності - це набір генів, спочатку придбаних з плазміди і тепер інтегрованих в бактеріальну хромосому. Він встановлює походження реплікації для хромосоми. Частина F - це «локомотив», який тягне хромосому в клітину-реципієнт, а решта - «кабуза».
У кишковій паличці приблизно один ген потрапляє через кожну секунду, що клітини залишаються разом. (Отже, для створення всього геному (4377 генів) потрібно близько 100 хв. Однак процес легко переривається, тому більш імовірно, що гени господаря, близькі за провідними генами F («локомотив»), зроблять це, ніж ті, що далі назад. «Кабуз» рідко робить його настільки не в змозі отримати повний фактор F, осередок одержувача продовжує бути «жіночим».
ДНК, яка робить його поперек, знаходить гомологічну область на жіночій хромосомі і замінює її (подвійним кросовером). Навмисно відокремлюючи клітини (в кухонному блендері) в різний час, можна визначити порядок і відносний інтервал генів. Таким чином можна скласти генетичну карту, еквівалентну генетичним картам еукаріотів. Однак тут інтервали карти - секунди, а не сантимооргани (см).
На малюнку 5.1.4 показаний механізм кон'югації клітин кишкової палички, де «чоловічому» не вистачає функціональних генів, необхідних для синтезу вітаміну біотину та амінокислоти метіоніну (Bio −, Met -), тому їх потрібно додавати до його культурального середовища. «Самка» має ці гени (Bio +, Met +), але має нефункціональні (мутантні) гени, які заважають їй синтезувати амінокислоти треонін та лейцин (Thr −, Leu -), тому вони повинні бути додається до його культурального середовища.
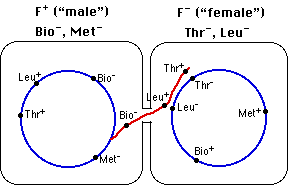
При спільному культивуванні деякі жіночі клітини отримують функціональні гени Thr і Leu від чоловічого донора. Подвійний кросовер дозволяє їм замінити нефункціональні алелі. Тепер клітини можуть рости на «мінімальному» середовищі, що містить тільки глюкозу і солі.
Трансдукція
Бактеріофаги - віруси, які заражають бактерії. У процесі складання нових вірусних частинок в них може бути включена деяка ДНК господаря. Головка віріона може вмістити лише стільки ДНК, тому ці віруси, поки ще здатні заразити нові клітини господаря і можуть бути не в змозі їх лізувати. Замість цього бактеріальний ген (або гени) автостопом може бути вставлений в ДНК нового господаря, замінивши ті, які вже є, і даючи господареві змінений фенотип. Це явище називається трансдукція.
Розуміння складних систем майже завжди доводиться чекати розгадки деталей якоїсь простішої системи. Ви можете відчути, що спроба з'ясувати, як один тип пневмокока може бути перетворений в інший, була надзвичайно спеціалізованим та езотеричним переслідуванням. Але Ейвері та його колеги усвідомили більш широке значення того, що вони спостерігали, і, з часом, і решта наукового світу. Вибравши працювати з чітко визначеною системою: перетворення R форм одного типу в S-форми іншого типу, ці дослідники зробили відкриття, яке зробило революцію в біології та медицині. Спроба зрозуміти роботу складних систем, спочатку зрозумівши роботу їх частин, називається редукціонізмом. Деякі вчені (і багато невчених) ставлять під сумнів значення редукціонізму. Вони виступають за цілісний підхід, що підкреслює роботу повної системи.
Однак запис говорить сама за себе. Від хмарочосів до місячних прогулянок, до комп'ютерних чіпів до досягнень сучасної медицини, прогрес приходить від першого розуміння властивостей частин, що складають ціле. Покійний Джордж Вальд, який отримав Нобелівську премію з фізіології 1967 року за відкриття молекулярної основи виявлення світла, колись переживав, що його робота надмірно спеціалізована - вивчаючи не зір, не око, не всю сітківку, навіть їх стрижні і конуси, а просто хімічні реакції їх родопсини. Але він зрозумів, що «це як би це було дуже вузьке вікно, через яке на відстані видно лише щілину світла. Коли хтось наближається, погляд стає все ширшим і ширшим, поки нарешті через це ж вікно не дивиться на Всесвіт. Я думаю, що так завжди йде в науці, тому що наука - це все одне ціле. Навряд чи має значення, куди входить людина, за умови, що можна підійти ближче...».
Значення генетичної рекомбінації бактерій
Трансформація, кон'югація та трансдукція були виявлені в лабораторії. Наскільки важливі ці механізми генетичної рекомбінації в природі? Завершення послідовності всього генома безлічі різних бактерій (і архей) говорить про те, що гени в минулому перейшли від одного виду до іншого. Це явище називається латеральним переносом генів (ЛГТ). Значному поширенню стійкості до декількох антибіотиків, можливо, сприяла передача генів резистентності всередині популяцій і навіть між видами. Багато бактерій мають ферменти, які дозволяють їм руйнувати чужорідні ДНК, що потрапляють в їх клітини. Здається, малоймовірно, що вони знадобляться, якби цього не відбувалося в природі. У будь-якому випадку, ці рестрикційні ферменти забезпечили інструменти, від яких залежать досягнення молекулярної біології та біотехнологічної промисловості.