4.12: Посередницький метаболізм
- Page ID
- 5485

Безпосереднім джерелом енергії для більшості клітин є глюкоза. Але глюкоза - не єдине паливо, від якого залежать клітини. Інші вуглеводи, жири та білки можуть бути в певних клітині або в певний час використовуватися як джерело АТФ. Складність механізму, за допомогою якого клітини використовують глюкозу, може змусити вас гаряче сподіватися, що подібно побудована система не потрібна для кожного виду палива. І дійсно це не так.
Одним з великих переваг поетапного окислення глюкози в СО 2 і Н 2 О є те, що кілька проміжних сполук, що утворюються в процесі, пов'язують метаболізм глюкози з метаболізмом інших молекул їжі.
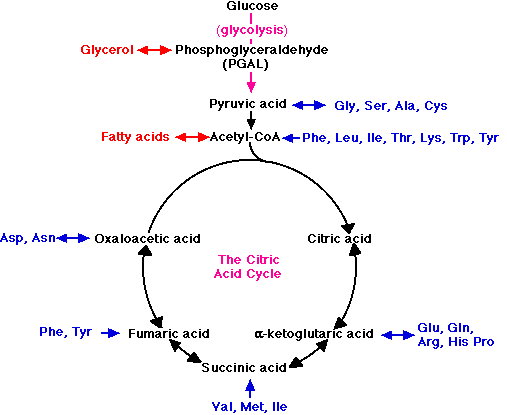
Наприклад, коли жири використовуються як паливо, гліцеринова частина молекули перетворюється в PGAL і потрапляє в гліколітичний шлях в цей момент. Жирні кислоти перетворюються в молекули ацетил-КоА і потрапляють в дихальний шлях для окислення в мітохондріях. Амінокислоти, що виділяються гідролізом білків, також можуть служити паливом.
- Спочатку видаляється азот, процес, який називається дезамінацією.
- Решта фрагменти потім потрапляють в дихальний шлях в декількох точках.
Для прикладів
- амінокислоти Gly, Ser, Ala, і Cys перетворюються в піровиноградну кислоту і потрапляють в мітохондрії, щоб бути дихати.
- Ацетил-КоА і кілька проміжних продуктів в циклі лимонної кислоти служать точками входу для інших фрагментів амінокислот (показані синім кольором).
Таким чином, ці зв'язки дозволяють дихати зайві жири та білки в раціоні. Не потрібен особливий механізм клітинного дихання тим тваринам, які значною мірою залежать від вживаних жирів (наприклад, багатьох птахів) або білків (наприклад, м'ясоїдних) для їх енергопостачання.
Значна частина білка, який ми споживаємо, в кінцевому підсумку перетворюється на глюкозу (процес, який називається глюконеогенезом), щоб забезпечити паливо для мозку та інших тканин. Хоча всі наші продукти в якійсь мірі взаємоконвертовані, вони не зовсім такі. Іншими словами, жодна їжа не може забезпечити всі наші анаболічні потреби. Ми дійсно можемо синтезувати багато жирів з глюкози, але певні ненасичені жири не можуть бути синтезовані і їх потрібно приймати безпосередньо в нашому раціоні. Це лінолева кислота, ліноленова кислота та арахідонова кислота. Всі ненасичені; тобто мають подвійні зв'язки. Хоча ми можемо синтезувати 11 амінокислот з попередників вуглеводів, ми повинні отримати 9 інших («незамінні амінокислоти») безпосередньо.
Багато точок, що з'єднують вуглеводний обмін з катаболізмом жирів і білків, служать двоходовими клапанами (позначені на малюнку двоголовими стрілками). Вони забезпечують точки входу не тільки для катаболізму (клітинного дихання) жирних кислот, гліцерину та амінокислот, але і для їх синтезу (анаболізму). Таким чином катаболічний розпад крохмалів може привести (через ацетил-КоА і PGAL) до синтезу жиру.
Фруктоза представляє особливу проблему
Фруктоза утворюється шляхом перетравлення дисахариду сахарози (звичайного столового цукру) в моносахариди глюкози і фруктози. І глюкоза, і фруктоза мають однакову емпіричну формулу (C 6 H 12 O 6) і однакову калорійність (686 ккал/моль). Але організм ставиться до них зовсім по-різному.
Глюкоза засвоюється і метаболізується всіма клітинами, щоб генерувати АТФ гліколізом та клітинним диханням, а надлишок глюкози переважно перетворюється на глікоген, а не жири. Фруктоза засвоюється тільки клітинами печінки, а надлишок фруктози перетворюється в жири (жирні кислоти і гліцерин). У США більшість безалкогольних напоїв та готових продуктів зараз підсолоджені кукурудзяним сиропом з високим вмістом фруктози (60% фруктози, 40% глюкози). Надмірне споживання цих продуктів цілком може бути пов'язане зі зростаючою поширеністю в США ожиріння та діабету 2 типу.