6.4: Картування обмежень
- Page ID
- 7413

Система обмеження/модифікації бактерій - це маломасштабна імунна система для захисту від зараження чужорідною ДНК.
Наприкінці 1960-х було виявлено, що кишкова паличка містить ферменти, які будуть метилювати специфічні нуклеотидні основи в ДНК.
· Різні штами кишкової палички містили різні типи цих метилаз
- Типові ділянки метилювання включають N 6 положення аденіну, положення N 4 цитозину або положення C 5 цитозину.
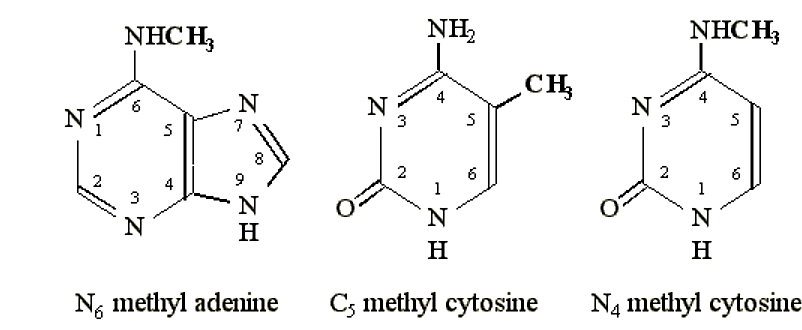.png)
Малюнок 6.4.1: Місця метилювання
- Крім того, лише дробовий відсоток основ був метильований (тобто, наприклад, не кожен аденін був метильований), і це відбувалося на дуже специфічних ділянках ДНК.
- Характерною особливістю ділянок метилювання, було те, що в них задіяні паліндромні послідовності ДНК.
- Ось приклад з конкретного штаму E. coli R1:
.png)
Малюнок 6.4.2: Паліндромна ДНК
(Специфічність метилази EcoR1. Рубін і Модрич, 1977)
- Крім того, що володіють певною метилазою, окремі штами бактерій також містили супутні специфічні ендонуклеазні дії.
- Ендонуклеази розщеплюються на місці розпізнавання метилювання або поблизу нього.

Малюнок 6.4.3: Розщеплення поблизу місця метилювання
- Однак ці специфічні нуклеази не розщеплювалися б при цих специфічних паліндромних послідовностях, якби ДНК була метильованою.
Таким чином, ця комбінація специфічної метилази та асоційованої ендонуклеази функціонувала як тип імунної системи для окремих штамів бактерій, захищаючи їх від зараження чужорідними ДНК (наприклад, вірусами).
- У штамі бактерій Ecor1 послідовність GAATTC буде метильована на внутрішній основі аденіну (метилазою EcoR1).
- Ендонуклеаза EcoR1 в межах одних і тих же бактерій не розщеплює метильовану ДНК.
- Тому чужорідна вірусна ДНК, яка не метилюється в послідовності «GAATTC», буде визнана «чужорідною» ДНК і буде розщеплена ендонуклеазою EcoR1.
- Розщеплення вірусної ДНК робить її нефункціональною.
Такі ендонуклеази називаються «рестрикційними ендонуклеазами», оскільки вони обмежують ДНК всередині клітини до «себе».
Поєднання рестрикційної ендонуклеази і метилази називається системою «обмеження-модифікація».
Оскільки різні штами та види бактерій мають потенційно різні системи R/M, їх характеристика зробила доступними сотні ендонуклеаз з різною послідовністю конкретних місць розщеплення.
- Вони є одним з основних інструментів сучасної молекулярної біології для маніпулювання та ідентифікації послідовностей ДНК.
- Рестрикційні ендонуклеази прийнято називати на честь бактерії, з якої вона була виділена.
|
Ім'я |
Джерело |
послідовність розпізнавання |
Коментарі |
|
Алу I |
Артробактер лютеус |
| 5'… A G C T … 3' 3'… T C G A … 5' | |
«Чотири різака». Листя тупими кінцями до ДНК. |
|
Бафа I |
Бактеріїди ламкі |
| 5'… C T A G … 3' 3'… G A T C … 5' | |
«Чотири різака». Листя 5' звис. |
|
Ніч I |
Нейссерія сінерея |
| C 5'… C C G G G … 3' 3'… G G C C C … 5' G | |
«П'ять різаків». Середнім підставою може бути як цитозин, так і гуанін. Листя 5' звис. Різні сайти розпізнавання можуть мати некомплементарні послідовності. |
|
Еко Р1 |
кишкова паличка |
| 5'… G A A T T C … 3' 3'… C T T A A G … 5' | |
«Шістка різак». Листя 5' звис. Поводиться як «чотири різака» (активність «зірки») у буфері з високим вмістом солі. $44 за 10000 одиниць. |
|
Хей II |
Гемофіл єгипетський |
| 5'… Pu G C G C Py … 3' 3'… Py C G C G Pu … 5' | |
«Шістка різак». Пу - будь-який пурин, Ру - будь-який піримідин. Листя 3' звис. |
|
Еко 109i |
кишкова паличка |
| 5'… Pu G G N C C Py … 3' 3'… Py C C N G G Pu … 5' | |
«Сім різак». Пу - будь-який пурин, Ру - будь-який піримідин, N - будь-яка основа. Листя 5' звис. Різні сайти розпізнавання можуть мати некомплементарні послідовності. |
|
Бгл I |
Глобігійна паличка |
| 5'… GCCN NNNNGGC … 3' 3'… CGGNNNN NCCG … 5' | |
«Шість різак з перерваним паліндромом». Листя 5' звис. Різні сайти розпізнавання можуть мати некомплементарні послідовності. |
|
Бас HI |
Бацила стеаротермофільна |
| 5'… G Pu C G Py C … 3' 3'… C Py G C Pu G … 5' | |
«Шістка різак». Різні сайти розпізнавання будуть доповнювати. |
|
Ат II |
ацетобактер ацетат |
| 5'… G A C G T C … 3' 3'… C T G C A G … 5' | |
«Шість різак» з 3' звисом. Та ж послідовність розпізнавання, що і Bsa HI, але різне положення розщеплення. |
|
Уд./хв I |
Бацила пумілус |
| 5'… C T G G A G N16 … 3' 3'… G A C C T C N14 … 5' | |
Непаліндромний, дистальний розщеплення. Листя 3' звис. $50 за 50 одиниць. |
|
Чи не я |
Нокардія отит |
| 5'… G C G G C C G C … 3' 3'… C G C C G G C G … 5' | |
«Різак вісімки». Листя 5' звис. |
|
БСМ I |
Бацила стеаротермофільна |
| 5'… G A A T G C N … 3' 3'… C T T A C G N … 5' | |
«Дивний». Листя 3' звис. |
- Корисність рестрикційних ендонуклеаз полягає в їх специфічності та частоті, з якою відбуваються місця їх розпізнавання в межах будь-якого заданого зразка ДНК.
- Якщо існує 25% ймовірності для конкретної бази на будь-якому даному сайті, то можна легко обчислити частоту, з якою будуть відбуватися різні ділянки обмеження ендонуклеази (0,25 п):
|
специфіка |
Приклад |
частота виникнення |
|
Чотири базові послідовності |
Алу I |
1 Alu сайт в кожні 256 баз (0.25 Kb) |
|
П'ять базових послідовностей |
Ніч I |
1 Nci I сайт в кожному 1024 бази (1.0 Kb) |
|
Шість базових послідовностей |
Декор I |
1 сайт eCor1 на кожні 4 096 баз (4.1 Kb) |
|
Сім базових послідовностей |
Еко 109i |
1 Ecoo109i сайт в кожних 16 384 базах (16.4 Кб) |
|
Вісім базових послідовностей |
Чи не я |
1 Не я сайт в кожному 65,536 баз (65.5 Kb) |
Таким чином, в середньому будь-яка дана ДНК буде містити сайт Alu I кожні 0.25 кілобази, тоді як сайт Not I відбувається один раз приблизно кожні 65.5 кілобази.
- Тому Not I є дуже корисним ферментом для виділення великих ділянок ДНК, як правило, в дослідженнях, пов'язаних з маніпуляціями геномної ДНК.
- Alu Я повинен був би перетравлювати зразок ДНК на безліч маленьких шматочків.
Асортимент фрагментів ДНК представляв би специфічний «відбиток пальця» конкретної ДНК, що перетравлюється. Різні ДНК не давали б однакову колекцію розмірів фрагментів. Таким чином, ДНК з різних джерел може бути або підібрана, або розрізнена на основі збірки фрагментів після рестрикційної ендонуклеази лікування. Вони називаються «Поліморфізми довжини фрагмента рестрикції», або RFLP. Цей простий аналіз використовується в різних аспектах молекулярної біології, а також правоохоронних органів та генеалогії. Наприклад, генетичні варіації, які відрізняють людей, також можуть призвести до меншої кількості або додаткового обмеження місць розпізнавання ендонуклеази.
Гелевий електрофорез ДНК
Найбільш поширеним гелевим електрофорезом твердої опорної матриці для молекул ДНК є
- агарози і
- акриламід.
Гелі ДНК агарози
Швидкість електрофоретичної міграції ДНК через агарозні гелі залежить від чотирьох основних параметрів:
1. Молекулярний розмір ДНК. Молекули лінійної дуплексної ДНК переміщаються через агарозні гелі зі швидкістю, яка обернено пропорційна журналу їх молекулярної маси.
М р α 1/лог (МВт)
Приклад: Порівняйте молекулярну масу та очікувану швидкість міграції:
|
Молекулярна маса (Da) |
Колод (Молец. Маса) |
1/журнал (Молец. Маса), |
|
100 000 |
5.0 |
0,20 |
|
50 000 |
4.7 |
0,21 |
|
10 000 |
4.0 |
0,25 |
|
5 000 |
3.7 |
0,27 |
|
1 000 |
3.0 |
0,33 |
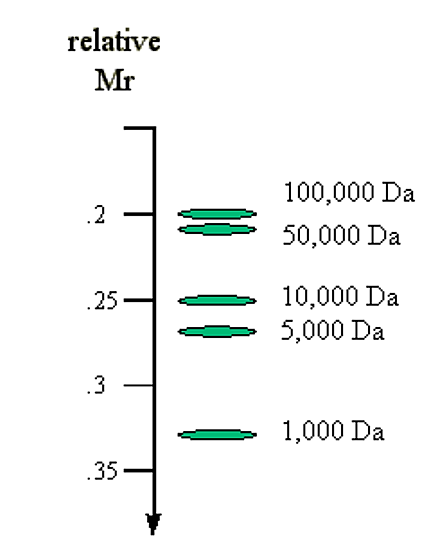.png)
Малюнок 6.4.4: Молекулярна маса і швидкість міграції
2. Концентрація агарози. Існує зворотна лінійна залежність між логарифмом електрофоретичної рухливості і концентрацією гелю.
|
Агароза (%) |
Діапазон поділу лінійної ДНК (в кілобазах) |
|
0.3 |
60 - 5 |
|
0.6 |
20 - 1 |
|
0.7 |
10 - 0,8 |
|
0.9 |
7 - 0,5 |
|
1.2 |
6 - 0,4 |
|
1.5 |
4 - 0,2 |
|
2.0 |
3 - 0,1 |
3. Конформація ДНК.
- замкнута кругова ДНК (Form-I) - типово перекручена (компактна)
- nicked circular (Форма-II) - нік розслабляє будь супернамотування
- лінійна ДНК (форма III)
Ці різні форми однієї і тієї ж ДНК мігрують з різною швидкістю через агарозний гель.
- Практично завжди лінійна форма (Form-III) мігрує з найповільнішою швидкістю з трьох форм
- Суперзгорнута ДНК (Form-I) зазвичай мігрує найшвидше
.png)
Малюнок 6.4.5: Форми ДНК
4. Прикладений напруга.
Інші деталі:
- Типове значення для запуску агарозного гелю становить 5 вольт на см (довжина гелю).
- Агарозні гелі зазвичай заливають і проводять горизонтально.
- Нарешті, ДНК, будучи кислою молекулою, мігрує до позитивно зарядженого електрода (катода). ДНК, природно, має постійне співвідношення заряду до маси, тому не потрібно додавати миючі засоби (як у випадку з білками)
.png)
Малюнок 6.4.6: Гелевий електрофорез
ДНК-акриламідні гелі
- Акриламідні гелі корисні для поділу дрібних фрагментів ДНК
- зазвичай олігонуклеотиди <100 пар основи.
- Ці гелі, як правило, мають низьку концентрацію акриламіду (<= 6%) і містять неіонний денатурующий агент сечовини (6M).
- Денатуруючий засіб перешкоджає утворенню вторинної структури в олігонуклеотидах і дозволяє відносно точно визначити молекулярну масу.
фарбування ДНК
- Найбільш зручним методом візуалізації ДНК в гелевому електрофорезі є фарбування флуоресцентним барвником бромід етидію.
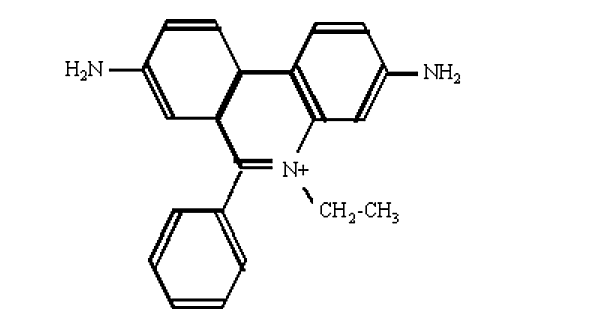.png)
Малюнок 6.4.7: Бромід етидію
- Ця сполука містить плоску групу, яка інтеркалюється між складені основами ДНК.
- Орієнтація і близькість етідію з укладеними основами змушує барвник проявляти підвищену муковитість порівняно з вільним барвником (у розчині).
- УФ випромінювання при 254 нм поглинається ДНК і передається пов'язаному барвнику.
- Енергія повторно випромінюється при 590 нм в червоно-помаранчевій області спектра.
- Бромід етідію зазвичай готують у вигляді вихідного розчину 10 мг/мл у воді, зберігають при кімнатній температурі і захищають від світла.
- Барвник зазвичай вводять в гель і працює буфер, або, навпаки, гель фарбують після прогону шляхом замочування в розчині броміду етідію (0,5 мкг/мл протягом 30 хв).
- Плівка візуалізується шляхом опромінення джерелом ультрафіолетового світла (тобто за допомогою транзилюмінатора) та фотографування поляроїдною плівкою.
- Звичайна чутливість виявлення краще 0,1 мкг ДНК.
Оскільки етидій є агентом інтеркаляції ДНК, він є потужним мутагеном. Включення етидію в ДНК живих організмів (тобто ви і я) може спричинити (небажані) мутації.
Поєднання рестрикційного ендонуклеазного травлення з гелевим електрофорезом ДНК: рестрикційне картування
Задана послідовність ДНК (наприклад, ген) матиме певну послідовність, а отже, специфічні ділянки рестрикції ендонуклеази
- Кількість і розташування таких ділянок є унікальною і передбачуваною властивістю для даної молекули ДНК.
- Картина фрагментації (тобто кількість і розмір фрагментів після рестрикції ендонуклеази перетравлення) може бути охарактеризована гелевим електрофорезом як тип «відбитка ДНК»
- Будь-які зміни в послідовності ДНК гена можуть призвести до ліквідації тих чи інших місць рестрикції, і навпаки, створювати нові:
.png)
Малюнок 6.4.8: Зміна сайтів обмежень
- Картина фрагмента гелевого електрофорезу для гена альбуміну людини і собаки буде характерно відрізнятися один від одного
- Зразок крові потенційно можна ідентифікувати як людину або собаку, спостерігаючи за поліморфізмом довжини рестрикційного фрагмента.
- Генетичні відмінності між особинами також можна виявити за допомогою аналізу поліморфізму довжини рестрикційного фрагмента.
.png)
Малюнок 6.4.9: Використання RFLP для визначення генетичних відмінностей