1.2: Структура ДНК та РНК
- Page ID
- 7414

Подвійна спіраль
ДНК (дезоксирибонуклеїнова кислота) і РНК (рибонуклеїнова кислота) складаються з двох різних класів азотовмісних підстав: пуринів і піримідинів. Найбільш часто зустрічаються пурини в ДНК - це аденін і гуанін:
.png)
Малюнок 1.2.1: Пурини
Найбільш часто зустрічаються піримідини в ДНК - це цитозин і тимін:
.png)
Малюнок 1.2.2: Пірамідини
РНК містить ті ж основи, що і ДНК, за винятком тиміну. Замість РНК містить піримідин урацил:
.png)
Малюнок 1.2.3: Тимін проти Урацилу
Аденін, гуанін, цитозин, тимін і урацил зазвичай скорочуються за допомогою однолітерних кодів A, G, C, T і U відповідно.
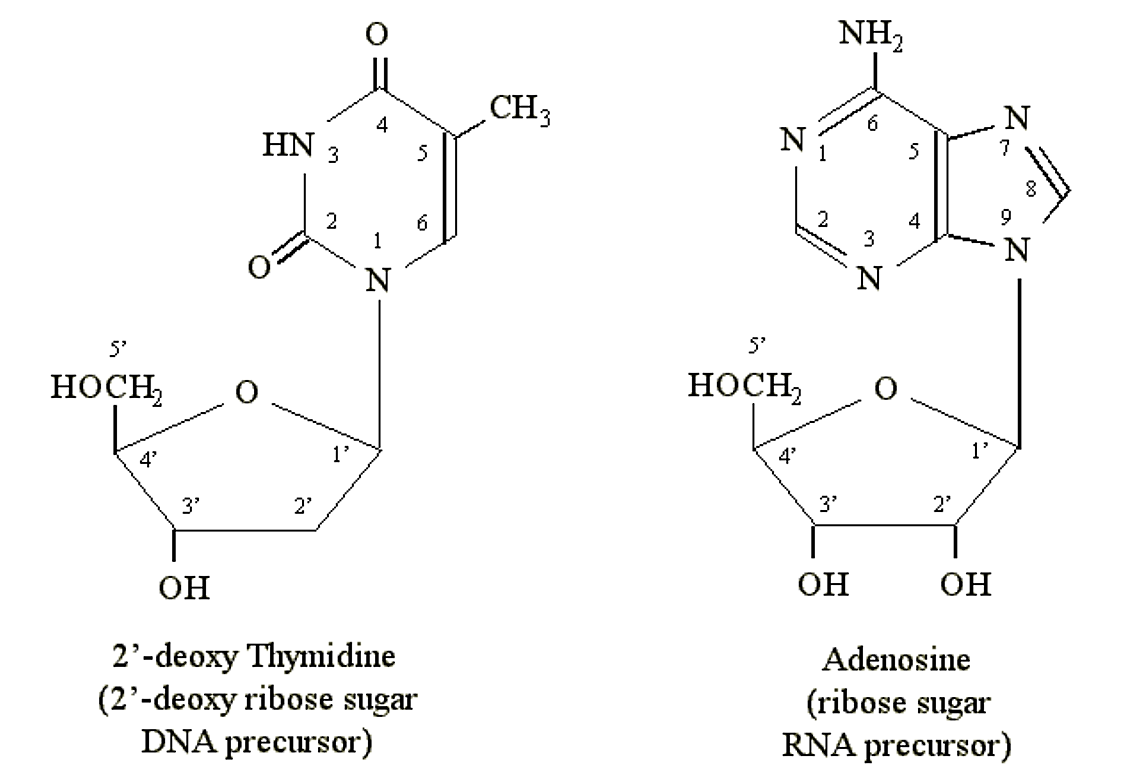
Пурини та піримідини можуть утворювати хімічні зв'язки з пентозними (5-вуглецевими) цукрами. Атоми вуглецю на цукрах позначаються 1', 2', 3', 4' і 5'. Це 1' вуглець цукру, який зв'язується з атомом азоту в положенні N1 піримідину або N9 пурину. Попередники ДНК містять пентозу дезоксирибозу. Попередники РНК містять пентозу рибозу (яка містить додаткову групу OH в положенні 2'):
.png)
Малюнок 1.2.4: Нуклеозиди
Перш ніж нуклеозид може стати частиною молекули ДНК або РНК, він повинен комплексуватися з фосфатною групою, щоб утворити нуклеотид (або дезоксирибонуклеотид, або рибонуклеотид). Нуклеотиди можуть мати 1, 2 або 3 фосфатні групи, наприклад, нуклеотиди аденозинмонофосфат (АМФ), аденозиддифосфат (АДФ) та аденозинтрифосфат (АТФ). Фосфатні групи прикріплені до 5' вуглецю рибозного цукру фрагмента. Починаючи з фосфатної групи, прикріпленої до 5' рибози вуглецю, вони маркуються a, b і g фосфат. Це трифосфатний нуклеотид, який включений в ДНК або РНК.
.png)
Малюнок 1.2.5: Нуклеотид
ДНК і РНК - це просто довгі полімери нуклеотидів, які називаються полінуклеотидами. До складу полімеру входить тільки фосфат. Він стає хімічно зв'язаним з 3' вуглецем цукрового фрагмента іншого нуклеотиду:
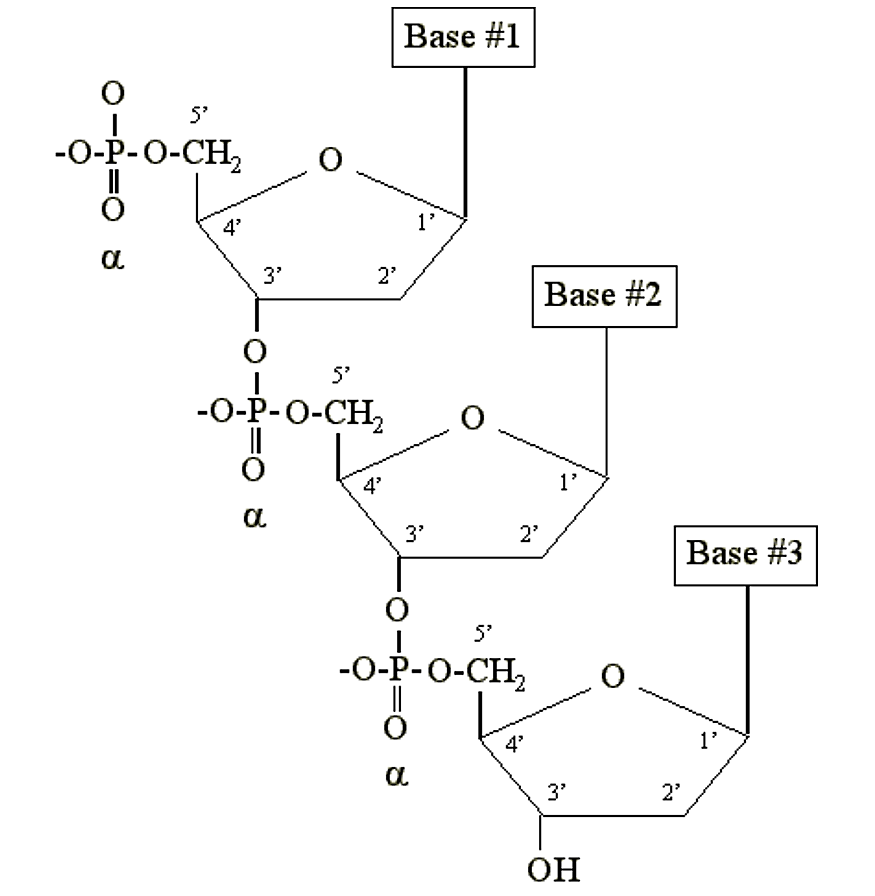.png)
.png)
Резюме термінів:
|
|
|
|
|
|
|
|---|---|---|---|---|---|
| Аденін | Аденозин | (Аденілова кислота) |
|
|
|
| Гуанін | гуанозин | (Гуанілова кислота) |
|
|
|
| Цитозин | Цитидин | (Цитидилова кислота) |
|
|
|
| тимін | тимідин | (Тимідилова кислота) |
|
|
|
| Урацил | Уридин | (Уридилова кислота) |
|
|
Яка структура ДНК? Як структура пов'язана з функцією?
1950-х
Була відома первинна хімічна структура полінуклеотидів (тобто зв'язок фосфату 3'-5').
1951 р. Чаргафф
Експеримент:
Візьміть ДНК з різних видів і гідролізуйте її для отримання окремих піримідинів і пуринів. Визначте відносні концентрації основ A, T, C і G.
Результат:
Хоча різні види мали однозначно різні співвідношення піримідинів або пуринів, відносні концентрації аденіну завжди дорівнювали концентрації тиміну, а гуанін дорівнював цитозину.
1950-х Р.Е. Франклін
Рентгенівські дифракційні дослідження волокон ДНК показали, що ДНК прийняла високо впорядковану спіральну структуру. Франклін зробив висновок, що дві або більше ланцюгів повинні обертатися навколо один одного, щоб утворити спіраль. Деякі основні розміри спіралі були розраховані за даними рентгенівської дифракції.
1953 Л. Полінг і Р.Б. Корі
Запропонуйте триланцюгову гвинтову структуру для ДНК з фосфатною кісткою в центрі та підставами зовні.
1953 Джей Д.Д. Ватсон і Ф.Крік
Виявлено розташування водневого зв'язку між моделями тиміну та аденіну основ, а також між цитозиновими та гуаніновими основами, які заповнені правилом Чаргаффа:
.png)
Малюнок 1.2.7: Склеювання правил Chargaff
Відзначимо, що пара «ТА» може накладати пару «ГК» з зв'язками на групи цукру в аналогічному зіставленні. У моделі «подвійної спіралі» Уотсона і Крика полінуклеотидні ланцюги взаємодіють, утворюючи подвійну спіраль з ланцюгами, що йдуть в протилежних напрямках. Підстави спрямовані до центру (і укладають один на одного), а цукрові хребти звернені на зовнішню сторону спіралі.
Модель Watson and Crick мала наступні фізичні розміри:
- 34 Å за спіральний повторення
- 10 пар основи за повторення (тобто за оборот спіралі)
- 3.4 Å міжбазова відстань укладання
- 20 Å діаметр для ширини гвинта
Фізичні характеристики моделі збігалися з тими, які були визначені рентгенівськими дифракційними дослідженнями Розалінд Франклін.
Наслідки моделі для генетичної інформації:
Документ Watson and Crick був вправою стислості (1 сторінка лише в Природі). Структура була настільки багата з підтекстом, що можна було написати зовсім небагато. Автори, однак, вирішили лише сказати: «Це не уникнуло нашого повідомлення, що конкретне сполучення, яке ми постулювали, негайно передбачає можливий механізм копіювання генетичного матеріалу».
- Якщо G завжди в парі з C, а T завжди в парі з A, то будь-яка пасмо може бути регенерована з додаткової інформації в іншій пасма.
- Основою комплементарності був водневий зв'язок, тобто нековалентні взаємодії, які можна було легко розірвати і повторно сформувати.
- Інформація, яку здійснювала ДНК, перебувала в межах унікальної базової послідовності ДНК.
- Із загального внутрішнього розташування баз, здавалося б, що подвійна спіраль повинна була б від'єднатися, щоб отримати доступ до інформації.
- Нерівнянне розташування цукрових фрагментів (див. Вище) припускало, що спіраль ДНК матиме велику канавку та незначну канавку.
Загальні позначення подвійної багатониткової ДНК:
.png)