1.24: Багаторазові ролі матеріального руху та дифузії в біології рослин
- Page ID
- 6956

Окрім необхідності в матеріалах, організмам потрібно переміщати матеріали: матеріали потрібно переміщати з місця їх придбання або синтезу в інші місця, де вони використовуються; матеріали, можливо, доведеться транспортувати до місць зберігання, а також витягувати з місць зберігання; хімічні сигнали можуть знадобитися перемістити з місце відчуття до місця реагування.
Матеріали переміщуються в організмах трьома основними шляхами, два з яких відбуваються і в неживих системах: (1) матеріали рухаються шляхом дифузії, що є наслідком того, що всі молекули при температурі вище абсолютного нуля (тобто всі молекули!) рухаються випадковим тепловим шляхом; (2) рідкі матеріали (тобто гази та рідини) рухаються масовим потоком, від високого тиску до низького до тих пір, поки існує відкритий шлях, який дозволяє рідини рухатися; (3) матеріали рухаються в результаті хімічних взаємодій, унікальних для біологічних утворень, так званих рухових білків, які можуть використовувати хімічну енергію для виконання фізичної роботи (тобто штовхання або витягування молекули, прикладання сили на відстань) або обертові двигуни джгутиків, які створюють обертальний рух, який «працює на» рух протона вниз електрохімічним градієнтом. Рухові білки є дуже значущими всередині клітин, де вони можуть виробляти цитоплазматичний потік - процес, який є значним для більших клітин, оскільки дифузія неефективна, за винятком дуже коротких відстаней. Більші клітини, знайдені в деяких водоростях, і особливо в коеноцитарних/сифонових клітині, сильно залежать від здатності рухових білків транспортувати матеріал всередині клітини. Але для цього курсу нас більше цікавить здатність більших багатоклітинних організмів, особливо рослин, переміщати матеріали всередині організму, процес, який здійснюється поєднанням дифузії та потоку тиску.
ТЕМИ
- Дифузія
- Проста, але недостатня модель
- Осмос - дифузія розчинників, включаючи воду
- Вплив тиску на дифузію
- Поєднання впливу чистоти і тиску
- Рослини і гриби використовують маніпуляції тиску і осмосу різними способами
- Структурно
- Зростання
- Охороняйте руху клітин
- Рухи листівки
- Міжміський транспорт
- У флоемі
- У ксилемі
- візерунки
- механізми
- проблема кавітації та компромісів в анатомії ксилеми
- рідкісні випадки ксилеми під тиском
Дифузія
Проста, але недостатня модель
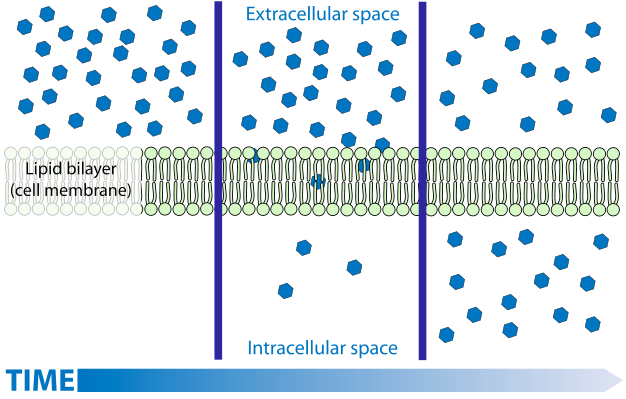
Дифузія - процес, знайомий більшості. Хоча дифузія газів і розчинених речовин легко зрозуміти спрощеними моделями, розуміння дифузії рідин, зокрема води, набагато складніше і часто заплутується неточним застосуванням термінології. Дифузія газів і розчинених речовин описується як спонтанний рух з областей більш високої концентрації в області меншої концентрації. Пояснення цього спонтанного процесу легко пов'язане з кінетичною теорією - молекули рухаються випадковим чином через теплову енергію. Як наслідок цього випадкового руху, відбувається чистий потік речовини з місць, де є більше молекул (тобто більш високі концентрації) в регіони, де менше молекул (менші концентрації). Якщо мати справу з сумішами (газ з більш ніж одним компонентом), кожен компонент буде рухатися незалежно від інших.
Дифузія рідкої води (осмос) НЕ завжди від «високої концентрації до низької»
Однак ця модель (пояснення) не легко застосовується до рідкої води з розчиненими речовинами або до розчинів взагалі, багато в чому тому, що ідея «концентрації» змінюється Концентрація не є настільки точним терміном, як можна подумати, він може бути виражений кількома різними способами (масова концентрація, концентрація числа, молярність, моляльність, молярність). У спрощеній моделі дифузії найбільш підходящим заходом буде (число) концентрація (кількість молекул на одиницю об'єму), оскільки випадковий рух переміщує молекули з місця, де вони мають більше молекул на одиницю об'єму, туди, де вони мають менше молекул на одиницю об'єму. Однак при розгляді розчинника (а не розчиненої речовини) концентрація числа не є точним предиктором дифузії. Для води та більшості розчинників кількість концентрації розчинника змінюється дуже мало, оскільки додаються розчинені речовини, але додавання розчинених речовин може мати істотний вплив на дифузію розчинника. І хоча обсяг води мало змінюється, коли додаються розчинені речовини, ступінь зміни змінюється з різними розчиненими речовинами, але вплив на дифузію води не контролюється конкретною доданою розчиненою речовиною, а лише від того, скільки розчиненої речовини (кількість частинок) було додано. Враховуйте, що в той час як додавання більшості розчинені речовини змушують воду трохи збільшуватися в обсязі (кількість концентрації знижується - така ж кількість молекул води зараз знаходиться в більшому обсязі), додавання деяких розчинених речовин може призвести до скорочення води (тобто концентрація кількості зростає - така ж кількість молекул води зараз знаходиться в меншому обсязі ). Якщо (кількість) концентрація води є тим, що спрямовує її рух, можна очікувати, що різні розчинені речовини матимуть різний вплив на дифузію рідини, залежно від того, наскільки вони спричинили зміну обсягу розчину (та концентрації кількості). І ви насправді очікуєте, що деякі рішення (ті, які спричиняють усадку води) матимуть воду дифузну з розчину в чисту воду. Такого ніколи не буває! Зберігаючи всі інші фактори постійними, вода завждидифується звідти, де вона чиста, в будь-який розчин, незалежно від розчиненого речовини. І при низьких концентраціях практично ніякого ефекту від конкретного розчину - всі вони мають однаковий вплив на дифузію незалежно від їх впливу на концентрацію кількості води. Це концентрація розчиненої речовини НЕ концентрація молекул води, які направляють дифузію: все інші фактори, що утримуються постійними, вода завжди дифундує звідки є менша концентрація розчиненої речовини, де є більш висока концентрація розчиненої речовини. З цього можна зробити висновок, що саме чистота води, а не її концентрація, керує дифузією. Це може здатися тонкою різницею, але насправді відображає деякі дуже глибокі особливості, пов'язані з законами термодинаміки. Чистота належить до ентропії, а ентропія, як відомо, «керує» спонтанними процесами. Більше того, наш «ментальний образ», тобто модель того, що викликає дифузію, не працює - молекули розчинника НЕ йдуть від того місця, де їх більше, туди, де їх менше, вони, як правило, йдуть від того місця, де вони чистіші до того, де вони менш чисті.
Вплив тиску на дифузію
Друга ключова причина того, що опис/модель, яка описує дифузію як відбуваютьсявід «високої концентрації до низької», є дефіцитним, полягає в тому, що він не враховує наслідки тиску. Тиск - це найвідоміша причина того, що рідини рухаються (вітер, потік води в трубах, кровотік у тварин), але ці рухи не є дифузією, це те, що називається «масовим потоком», рухом, який залежить лише від перепадів тиску, і рух, який буде відбуватися щоразу, коли є перепади тиску та відкритий шлях для потоку рідини. Але коли масовий потік неможливий (тому що немає «відкритого шляху») тиск також може впливати на дифузійний рух: вода буде дифундувати з областей високого тиску в області низького тиску. Особливо це важливо для клітин, що володіютьяк клітинною мембраною, так і клітинною стінкою. Хоча мембрана дозволяє існувати різні чистоти розчинника (води) всередині проти зовнішньої клітини, стінка дозволяє існувати різний тиск всередині проти зовнішньої клітини, і як чистота, так і тиск важливі для диктування дифузії води.
Поєднання впливу чистоти і тиску
За інших рівних умов вода переміщається дифузією з областей більш високого тиску в області нижчого тиску, а також з областей підвищеної чистоти в райони низької чистоти. Ці два фактори можуть «збалансувати» один одного, і можна не мати дифузії між областю низької чистоти та високого тиску, пов'язаною з областю низького тиску та високої чистоти. Дивлячись на це по-іншому, якщо у вас вода низької чистоти обмежена в жорсткій ємності (тобто осередку з клітинною стінкою) і вона поміщена в чисту воду, вода буде рухатися в осередок, підвищуючи тиск в осередку. Зрештою, тиск буде досягнуто там, де більше немає дифузії в клітину. У цей момент різниці тиску між внутрішньою та зовнішньою частиною відповідають різниці чистоти між внутрішньою та зовнішньою стороною.
На жаль, не існує легко концептуалізованої моделі дифузії рідкої води, оскільки існує для дифузії газів та розчинених речовин. Сувора модель дифузії рідкої води вимагає застосування понять з термодинамічного параметра, який називається водним потенціалом. Основна ідея відносно проста: осмос (дифузія рідкої води) - це спонтанний процес, і будь-який спонтанний процес повинен призвести до зменшення кількості енергії, доступної для роботи («вільна енергія»). Як правило, на вільну енергію води (її водний потенціал) впливають два ключові фактори: тиск (що збільшує її водний потенціал) та наявність розчинених речовин (що зменшує її водний потенціал). Рідка вода дифундує від областей з високим потенціалом води в райони з низьким потенціалом води так само, як тепло тече від теплих районів до холодних районів.
Для всіх організмів, і зокрема тих, хто не має клітинної стінки, клітинна мембрана та клітинна активність дозволяють розвиватися різниці концентрації розчинених речовин між внутрішньою та зовнішньою частинами клітини, що призводить до відмінностей у чистоті води між внутрішньою частиною клітини та зовнішньою. Коли розвиваються відмінності в чистоті, вода буде надходити всередину або назовні шляхом дифузії. Поки відмінності в чистоті невеликі, цей рух може усунути відмінності в чистоті, зробивши внутрішню частину менш чистою (якщо вода витікає) або більш чистою (якщо вода тече). Однак такий рух води також призведе до зміни обсягу клітини, і якщо клітина набрякає або скорочується занадто сильно, це може завдати непоправної шкоди мембрані, тим самим руйнуючи функціонування клітин. Як наслідок, організми без клітинної стінки повинні або жити в районах, де чистота води подібна до того, що знаходиться всередині їх клітин, або вони повинні мати структури/механізми, що зменшують дифузію та/або мати здатність або усунути воду, яка дифузує в (наприклад, скорочувальна вакуоль Парамецій) або придбати воду, щоб замінити втрату дифузією (як правило, це досягається шляхом придбання «солоної» води та усунення солей).

Для клітин зі стінкою (наприклад, рослин і грибів) стіна дозволяє нове «рішення» для життя в районах, де клітина більш концентрована в розчинених речовині, ніж зовнішнє середовище (це нормальна ситуація для більшості неморських середовищ існування: прісноводних озер і струмків та наземних середовищ існування, де знаходяться організми занурений частково або повністю в грунт, вода якого, як правило, досить чиста, тобто з невеликою кількістю розчинених речовин). Для цих організмів жорстка стінка дозволяє тиску збільшуватися в міру надходження води в клітини. Цей тиск діє для зменшення внутрішньої дифузії води і в кінцевому підсумку досягається динамічна рівновага, де високий тиск і низька чистота всередині клітини врівноважують нижчий тиск і більш високу чистоту поза клітиною. Вода рухається (дифузно, тобто рухається по осмосу) всередину і назовні з однаковою швидкістю.
Рослини та гриби використовують «осмотичні системи» різними способами
Структурно
Вода може використовуватися як «будівельні блоки», коли вона обмежена структурою, яка не буде розширюватися. Живі клітини, при поєднанні клітинної мембрани і клітинної стінки, структурно міцні і рослини і гриби використовують їх для формування жорстких структур, здатних протистояти гравітаційним і вітровим силам (також розглянуто в главі 3). Докази структурної важливості води надходять із спостереження за в'яненням: якщо рослини позбавлені джерела води, щоб замінити втрачену випаровуванням, вони втрачають структурну цілісність. Деякі рослини та гриби виробляють деякі структури (наприклад, дерева, гриби-брекети), які не руйнуються, коли вони позбавлені води, але для багатьох рослин доступ до води має важливе значення для «стоячи», оскільки саме тиск клітин забезпечує жорсткість. Центральним елементом цієї здатності є клітинна стінка, яка має високу міцність на розрив і протистоїть розширеннюі, отже, дозволяє забезпечити тиск.
Зростання
Клітинний ріст відбувається тоді, коли внутрішній тиск перевищує міцність клітинної стінки і тим самим поступається тиску всередині неї. Органи (грибкові нитки, коріння і пагони) ростуть в результаті розширення окремих клітин і внутрішній тиск не тільки змушений виштовхувати клітинну стінку, але і, можливо, доведеться відштовхувати (стискати) грунт на своєму шляху. Таким чином, для рослин (як коренів, так і іноді пагонів) і грибків зростання може зажадати вироблення значної сили. Сила для живлення цього зростання походить від дифузії води (осмосу) і дуже значні сили можуть бути створені в результаті руху води вниз по її градієнту водного потенціалу. Тиск 2-4 бари (= 0,2-0,4 МПа [мегапаскаль] = 2-4 атмосфери тиску = 30-60 фунтів на квадратний дюйм) є поширеним явищем і може бути досить ефективним, як міг зрозумів кожен, хто спостерігав кульбаба, що піднімається через тротуар.
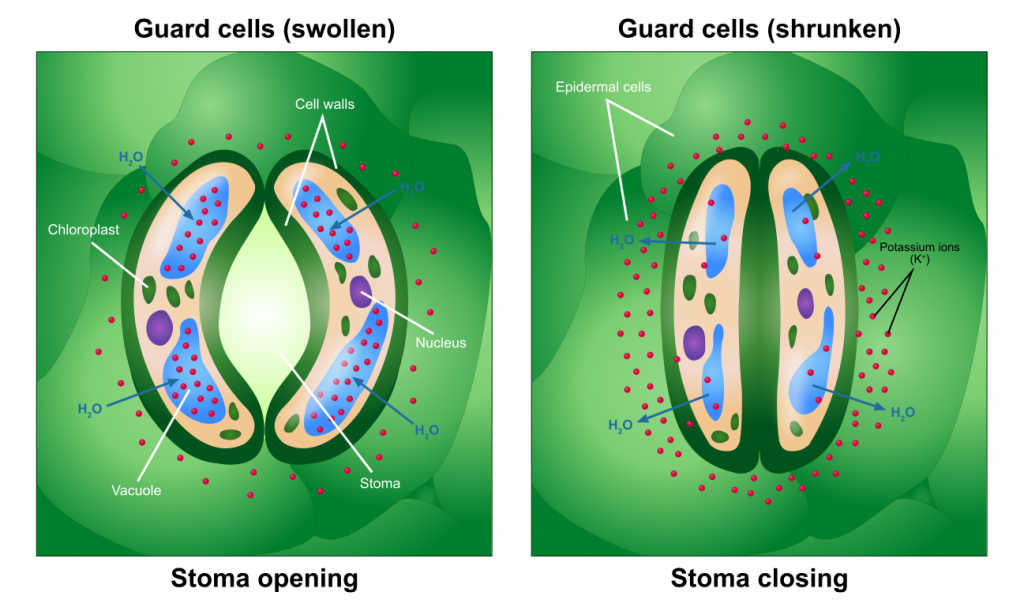
Охоронні клітини і стомати
Відкриття і закриття стоматів відбувається в результаті зміни тиску спеціалізованих клітин, охоронних клітин, які оточують пори. Герметизація захисних клітин в результаті накопичення розчинених речовин і подальшої дифузії води в охоронні клітини змушує клітини набрякати і утворювати між ними отвір (стомат). Зменшення розчинених речовин у захисних клітинапризведе до руху води з клітини, що призведе до падіння тиску і, отже, устьинного закриття. Тригери, які стимулюють захисні клітини накопичувати або втрачати розчинені речовини, були широко вивчені і включають світло та концентрацію вуглекислого газу. Точний контроль може бути не однаковим для всіх видів. Принаймні деякі рослини здатні регулювати внутрішній рівень вуглекислого газу в «заданій точці», що дозволяє фотосинтезу протікати з невеликим гальмуванням через брак вуглекислого газу, одночасно мінімізуючи кількість води, втраченої через транспірацію.
Рух листівки
Подібно до дії охоронних клітин, ряд рослин мають листя або листівки, які рухаються у відповідь на екологічні сигнали, такі як світло, дотик та посуха, в результаті чого утворюються листя або листівки, орієнтація яких змінюється залежно від обставин. Поширеним прикладом є «рухи сну», де листя горизонтальні вдень і вертикальні вночі. Ці рухи є результатом зміни тиску «пульвінарних» клітин, розташованих в точках повороту. Відносно невеликі зміни в розмірах цих клітин задіяні в результаті їх розташування і можуть спричинити суттєві змінистановища задіяних органів. (див. http://www.youtube.com/watch?v=U-PK13JEgk8 нижче)
Дивитися

Елемент YouTube був виключений з цієї версії тексту. Ви можете переглянути його онлайн тут: pb.libretexts.org/sdt34b/? p=302

Міжміський транспорт
Флоем
Обидві транспортні системи в рослині, транспорт ксилеми та флоем, працюють в результаті перепадів тиску, що створюються в «трубах», що знаходяться в цих тканині. Перепади тиску створюються в результаті дифузії води. У флоемі тиск створюється в результаті додавання розчинених речовин (цукрів) до клітин труб, процесу, відомого як флоемне навантаження, який відбувається в регіонах рослини, які називаються «джерельними» районами. Внаслідок флоемного навантаження вода надходить в ситові трубчасті елементи вихідної області, а тиск збільшується, запускаючи потік в трубах. Різниця тиску та потік підтримуються, оскільки розчинені речовини не просто завантажуються в «джерельні» ділянки, вони також видаляються в місцях «раковини», місцях, до яких транспортуються розчинні речовини; таким чином, у флоемі відбувається безперервний градієнт тиску, від джерел, що забезпечують цукри, до раковин, які споживають або запасних цукрів. Точні місця, які служать джерелами або раковинами, можуть змінюватися залежно від того, чи є місце вироблення цукрів (сахарозу) або їх споживання. Рух флоема може бути вгору по рослині (наприклад, від місць зберігання в корені, щоб стріляти верхівковими меристемами) або вниз по рослині (наприклад, від фотосинтезуючих листя до зберігання сайти в корені). Завантаження сахарози вимагає метаболічної енергії, оскільки АТФ використовується для переміщення сахарози звідки вона менш концентрована туди, де вона більш концентрована. Як і у випадку з охоронними клітинами, рух води в ситові трубки є пасивним після додавання розчинених речовин. Оскільки це масовий потік, не тільки сахароза, а будь-яка розчинена речовина, яка знаходиться в ситовій трубці, буде транспортуватися до раковини. Найбільш поширеними з цих інших розчинених речовин є амінокислоти, але інші азотовмісні сполуки транспортуються у флоемі разом з деякими мінеральними елементами (наприклад, K +, Mg 2 +, Ca 2 +)
Ксилема

Водний транспорт в ксилемі також є результатом «різниці тиску», але це насправді відмінності в напрузі, а не різниці в тиску. У той час як тиск стискає рідини, напруга витягує рідини, так само, як натягування на струну надає силу, яка діє, щоб розірвати молекули струни. Дивно, але в певних ситуаціях вода має значну міцність на розрив і дійсно може бути витягнута. Клітинні стінки всіх клітин в листі вливаються і покриті водою через адгезію води до клітинних стінок і згуртованості води з собою. Коли вода залишає лист шляхом випаровування, залишок води «розтягується», ставиться під напругу, тому що залишився менший обсяг води покриває той же початковий об'єм осередків. Ця напруга передається воді в провідних осередках ксилеми і створює «різницю тиску» (насправді різниця натягу), яка може тягнути воду вгору (неживі) клітини труби. Те, що, як правило, рухає рух води вгору по трубах (trach ei ds і судини) - це напруга, створена, оскільки вода втрачається через транспірацію. Однак потік води вгору по ксилемі також може відбуватися, навіть якщо транспірації мало до тих пір, поки відбувається зростання: дифузія води в розширюються клітини може створити напругу, щоб витягнути воду вгору рослин навесні, коли рослинам не вистачає листя і швидкість транспірації дуже низька.
Втрата води з листа проста дифузія: водяна пара у високих концентраціях (висока вологість повітря всередині листа) дифундує через відкриті продихи туди, де вологість нижча за межами листа. Вологість повітря всередині листа підтримується тому, що рідка вода в клітинних стінках клітин мезофілів випаровується і замінює втрачену воду. Напруженість, породжена втратою води, призводить до того, що трахеїди та судини злегка стискаються, оскільки тиск зовні є «нормальним» (одна атмосфера), але тиск всередині нижчий. Зверніть увагу, що це на відміну від ситуації в живих клітинам мезофілів, які знаходяться під тиском, оскільки вони мають мембрану, яка дозволяє їм концентрувати розчинені речовини. Якщо напруга в товщі води стає занадто великим, кавітація виникає, як бульбашки повітря утворюються при порушенні товщі води або при відриві води з боків трахеїди або посудини. У будь-якому випадку клітина «кавітаційна» і більше не корисна для водного транспорту.
Як у ситуаціях, керованих транспірацією, так і у транспорті флоемів, існує дифузійний рух на короткі відстані, що створює різницю тиску, яка може призвести до руху на великі відстані. Як в ксилемі, так і в флоемі рух всередині труб НЕ є дифузією, це набагато більш звичний процес, який називається об'ємним потоком. Об'ємний потік є набагато більш ефективним засобом транспортування матеріалів на великі відстані, ніж дифузія, яка ефективна лише на дуже малих відстанях (десяті частки мм для рідин). Об'ємний потік блокується клітинними мембранами і перешкоджається, але не перешкоджає клітинним стінкам. Ями, знайдені в клітинних стінках трахеїдів і елементів труб судин, забезпечують відносно низький шлях опору для переміщення води між сусідніми клітинами. Вода тече ще легше через перфораційні пластини судин, оскільки вони повністю відкриті. У (живих) провідних клітині флоеми плазмодезмати з'єднують окремі клітини і флоемний сік (який по суті є цитоплазмою, в якій відсутні органели) об'ємний потік відбувається від клітини до клітини через плазмодесмати. Об'ємний потік також значний у ґрунті, де є проходи для протікання води, і де як гравітація, так і «тяга» при русі в рослини можуть створювати перепади тиску.
Тиск і напруженість, виявлені в судинній тканині, відображають ці механізми. Якщо пробивається ситова трубка, флоемний сік витікає, оскільки тиск всередині клітини більший, ніж атмосферний, так само, як ви будете кровоточити, якщо ваша шкіра розірвана. За більшості обставин, якщо в трахеїд або посудину проникає вода НЕ витікає назовні, швидше за надходить повітря, відображаючи той факт, що вода всередині трахеїда/судна перебувала під напругою. Насправді, якщо вимірювати об'єм трахеїди/судини в міру розвитку напруженості, він трохи зменшується через стиснення ззовні. Через це стовбури дерев демонструють помітне зменшення окружності протягом дня, оскільки транспірація та напруженість збільшуються, і вони відскакують протягом ночі, коли напруга знімається, а рослина регідратується під час незначного або відсутнього транспірації.
У рідкісних випадках вода в ксилемі знаходиться під тиском.
Ця умова описується як «кореневий тиск» і виникає лише за певних обставин. «Кореневий тиск» - це «кровотеча» (ексудація) з обезголовленого стебла. За цих же особливих умов, якщо проколоти окрему посудину ксилеми або трахеїду, він також кровоточить, на відміну від більш нормальної ситуації, описаної вище. Тиск коренів виникає лише в тому випадку, якщо грунт вологий, коріння активно ростуть, а транспірація низька (вночі або коли немає листя). У цих умовах розчинні речовини (мінеральні іони) накопичуються в ксилемі кореня через дії кореневих клітин, і тому, що ендодерміс колективно поводиться як мембрана і є бар'єром, який запобігає витіканню розчинів, накопичених у кореневій ксилемі, назад з тканини ксилеми. Отже, як жива окрема рослинна клітина, яка може тиснути, оскільки клітинна мембрана дозволяє накопичувати розчинені речовини, вся коренева ксилема може накопичувати іони та тиснути. Це явище рідкісне, як тому, що коріння, як правило, недостатньо активні для накопичення достатньої кількості іонів у тканині ксилеми, а також тому, що витяг зверху (створений транспірацією або зростанням) запобігає накопиченню тиску.
Остаточна ситуація, коли ксилема знаходиться під тиском, - це та, яка змушує сік витікати (виходити) у стовбурах клена наприкінці зими та ранньою весною. «Постукування», вставляючи циліндр, у ксилему, призводить до кровотечі соку, який можна збирати, концентрувати та використовувати як джерело цукру. Кленовий сокорух НЕ вимагає кореневої активності — його можна спостерігати в стеблах, видалених з кореневої системи. Кленовий сік дійсно вимагає циклів заморожування/відтавання, оскільки вони так чи інакше дозволяють ксилемі стати під тиском. Цукор, що міститься в соку ксилеми, надходять з ксилемних променів, крохмаль яких перетворюється в сахарозу наприкінці зими. Чому тільки клени та деякі інші породи дерев демонструють таку поведінку, як правило, приписують аспектам їх анатомії деревини. Більшість порід дерев не тиснуть під впливом циклів заморожування/відтавання.
Хоча тканина флоема проникає при постукуванні клена, і хоча тканина флоема знаходиться під тиском і транспортує сахарозу, надзвичайно мало соку, зібраного з кленів, виходить з флоеми. Живі клітини (елементи ситової трубки) флоемної тканини здатні швидко закупорювати отвори, щоб запобігти «кровотечі» і, як наслідок, втрати цукру. Кленові дерева, які постукують, кровоточать з тканини ксилеми протягом декількох місяців, якщо вони піддаються належним умовам і до тих пір, поки немає «тяги» з верхівки рослини, викликаного ростом або випаровуванням з листя. Кленовий сік тече, коли листя немає і зростання не відбувається. Якби хтось міг потрапити в тканину флоема, було б виявлено, що концентрація цукру набагато, набагато вища (порівнянна з кленовим сиропом), ніж кленовий сік, солодкість якого ледь виявляється людьми, поки вона не концентрується.