1.5: Клітинна структура в неживому житті
- Page ID
- 7112

Відкриття першої клітини часто приписують Роберту Гуку в середині 1 7-го століття. Але люди спостерігали клітини і раніше (риб'ячі ікри, ікри жаб, яйцеклітини птахів). Важливим у спостереженні Гука було те, що він зазначив, що організми, принаймні деякі з них, складаються з менших сутностей, які він називав клітинами. Він зміг побачити їх частково тому, що він використовував нещодавно розроблений мікроскоп. Але те, що також зробило клітини видимими, це те, що вони були рослинними клітинами, клітинами, які не тільки більші за клітини більшості тварин, але також мають клітинні стінки, які окреслюють кожну клітинку і є жорсткими і не руйнуються, коли матеріал розрізається на тонкі ділянки. Матеріал, на який він дивився, була пробка, зовнішня кора дерева, і те, що він спостерігав, - це залишки клітин, які більше не були живими; їх існування збереглося через їх товстих і стійких клітинних стінок (рис. 1).

Хоча спостереження Гука проілюстрували, що принаймні деякі частини деяких організмів складаються з менших одиниць, іон генералізат, що всі організми складалися з однієї до багатьох «фундаментальних одиниць», клітин, не повинен був бути повністю реалізований ще 200 років, коли була запропонована та відстоюється «теорія клітин». Хоча люди були дуже обізнані про риб'ячі ікри та жаб'ячі ікри, вони не розуміли, що коли ікринки перетворилися на пуголовків, матеріал спочатку був розділений на багато, багато дрібних одиниць, кожна з однаковою основною конструкцією. Згодом ці підрозділи диверсифікувалися один від одного і трансформувалися в групи, щоб служити певним ролям. Т вони влаштовані певним чином для отримання складного цілого, зародка жаби. Подумайте про те, щоб зробити замки з лего. Подібна трансформація відбувається в міру перетворення заплідненої яйцеклітини рослин в ембріон рослини, хоча яйцеклітина рослин (близько 20 мкм) в 100 разів менше, ніж у жаб і приблизно в п'ять разів менше, ніж у людей.
Однією з великих тем біології є єдність і різноманітність життя: як організми одночасно можуть бути такими подібними і такими різними. Ця ж концепція стосується і клітин і є основоположною для «теорії клітин»: живі істоти складаються з клітин, основна організація, хімічний склад та організація яких схожі; але ці клітини також різноманітні за формою та функцією, і різноманітність спостерігається як між різними організмами (рослинні клітини відрізняються від клітин тварин), але також всередині (багатоклітинних) організмів.
Різноманітність клітин в організмі є єдиним, і для деяких людей єдиним критерієм багатоклітинного організму не просто те, що вони складаються з декількох клітин, але що принаймні деякі клітини відрізняються один від одного. Перш ніж описувати різноманітність типів клітин, виявлених у деяких організмах, які ми охоплюємо, ми розглянемо додаткові критерії, які можуть бути використані для визначення багатоклітинності.
ТЕМИ
- Багатоклітинність
- Клітинна структура грибів
- Хітриди
- Коеноцитарні, нитчасті гриби — хлібні цвілі та гломеромікоти
- Септат гриби — Клуб і мішок гриби
- Клітинна структура у формі води
- Будова клітин у водоростей
- Бурі водорості
- Червоні водорості
- Зелені водорості
Багатоклітинність

Чи може одноклітинний організм бути багатоклітинним? Можна подумати, що це абсурд, але це може бути правдою, залежно від визначень. Показовим випадком є Апікомплекс, група, до якої входить причинний організм малярії, Плазмодій. Члени цієї групи завжди одноклітинні, але мають кілька типів клітин (рис. 2), клітини, які виглядають по-різному, поводяться по-різному і живуть в різних місцях. Більшість біологів не вважали б Апікомплекс багатоклітинним і вважали б його біологічною сутністю з декількома формами, але в сенсі розвитку це ставить те саме питання, яке ставить багатоклітинність: якщо одна клітина генетично ідентична іншій клітині, як вони можуть в кінцевому підсумку виглядати? і поводитися по-іншому?
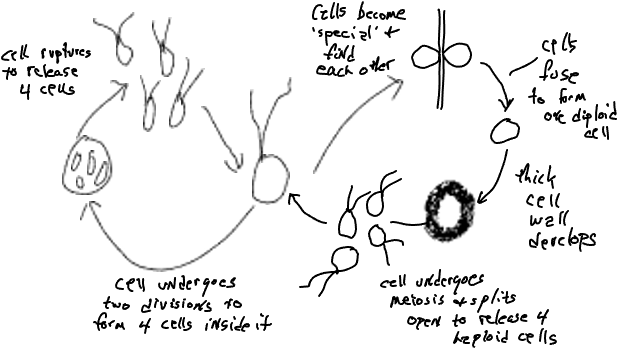
Якщо уважно вивчити життєвий цикл плазмодію, то відзначимо, що деяка частина клітинного різноманіття пов'язана з розмноженням і статевим процесом, тема, яка буде розглянута більш детально пізніше, а поки просто відзначимо, що більшість організмів виробляють. різні клітини, які пов'язані з розмноженням і/або статтю. Хламідомонада, одноклітинна зелена водорость (рис. 3), виробляє бі-джгутикові клітини, які виглядають і поводяться однаково, за винятком: (1) деякі клітини виглядають по-різному, оскільки вони втрачають свої джгутики і діляться двічі, щоб виробляти чотири клітини, перебуваючи в межах батьківської клітини. Зрештою батьківська клітинна стінка перетравлюється, і чотири типові на вигляд клітини звільняються, (2) деякі клітини виглядають нормально, але є «особливими», функціонально різними, оскільки вони здатні зливатися з іншими «спеціальними клітинами». Клітини стають «особливими» через зміну умов, наприклад, поживні речовини стають менш доступними. (3) клітина, утворена злиттям двох спеціальних клітин, розвивається в дуже різну на вигляд клітину, одну з яких не має джгутиків і товстої клітинної стінки. Він також функціонує по-різному, будучи метаболічно неактивним. Врешті-решт він стає активним і ділиться, утворюючи чотири типові виглядають (бі-джгутикові) дочірні клітини, які вивільняються після того, як клітинна стінка перетравлюється і ослаблена. Три атипові типи клітин пов'язані з статтю і процес мейозу (права сторона наведеної вище діаграми) буде розглянуто більш детально пізніше, разом з розглядом відтворення (ліва сторона діаграми).
Очевидно, що дуже мало хто вважає одноклітинні організми, такі як Plasmodium або Chlamydomonas, багатоклітинними, незважаючи на те, що вони можуть проявляти кілька типів клітин. Справа в тому, що володіння різними або різними поведінковими клітинами (наприклад, яйцеклітиною, спермою, зиготою, спорою), які пов'язані з репродуктивним або статевим процесом, недостатньо, щоб позначити організм як багатоклітинний. Це банально вірно для одноклітинних організмів, але також стосується колоніальних організмів, які складаються з декількох клітин, які майже всі однакові, і є причиною відмінності між «простою багатоклітинністю» та «складною багатоклітинністю».

Розглянемо нитчасті зелені водорості Ulothrix, (рис. 4), які існують у вигляді рядків клітин, більшість з яких виглядають однаково (див. Центральну частину малюнка вище, позначену «a»), але іноді деякі клітини перетворюються (тобто розвиваються), утворюючи рухливі клітини, які мають чотири джгутики ('c' на схемі) і звільняються від нитки розжарювання. Ці клітини можуть встановлювати нові нитки, прикріпивши до підкладки та діливши для отримання нової нитки ('d'). Інші клітини трансформуються і виробляють, а пізніше звільняють клітини лише з двома джгутиками ('f'). Ці клітини повинні знайти іншу біджгутикову клітину, злитися з нею ('g'), потім пройти мейоз, утворюючи чотири чотирибігучі клітини ('h'), які можуть прикріпитися до субстрату і утворювати нові нитки.
Інший приклад простої багатоклітинності з різноманітними типами клітин, пов'язаними з розмноженням і статтю, знаходиться в Oedogonium, (рис. 5) ще одна нитчаста зелена водорость. Більшість осередків циліндричні з довжиною приблизно в п'ять разів більше їх ширини. Але є також яйцеклітини, які коротші і опуклі посередині і набагато коротші клітини, що виробляють сперму, з довжиною однаковою або коротшою, ніж їх ширина. Обидва ці типи клітин розвивають отвори в їх стінках, що дозволяє коротким клітинам вивільняти рухливу сперму і дозволяє спермі проникати в яйцеклітини, що призводить до вироблення клітини з генетичною інформацією від двох батьків. Це переростає в сплячу спору, яка з часом виділяється з нитки. Додатковий тип клітин може розвиватися з нормальних клітин, оскільки вони розвивають джгутики і в кінцевому підсумку звільняються з нитки як клітини, здатні утворювати нових особин.

Ці приклади демонструють загальні типи клітин, пов'язані з статтю та розмноженням та виявлені в організмах з колоніальним зростанням (демонструючи «просту багатоклітинність»): (1) клітини, здатні зливатися між собою, називаються гаметами, які іноді диференціюються на яйцеклітину та сперму, (2) зигота, клітина, утворена злиття гамет, (3) зооспор, джгутикових клітин, спеціалізованих для рухливості (розсіювання в просторі) (наприклад, клітина, позначена «c» на діаграмі Ulothrix), (4) неактивні, сплячі клітини, які дозволяють розсіюватися в часі. Взагалі, вони називаються спорами (але врахуйте, що зооспори НЕ є неактивними). Ці неактивні клітини називаються ендоспорами у бактерій і акінетами в ціанобактеріях. У ряді організмів (наприклад, хламідомонади, оедогоній, хлібна цвіль) спори розвиваються з зигот і називаються зигоспорами. Ми побачимо в грибах ряд різних суперечок.

Повторюючись, жоден із розглянутих прикладів не вважатиметься багатоклітинним, хоча вони виробляють кілька клітин, деякі з яких структурно та функціонально відрізняються від інших. Які додаткові функції сприяють справі «справжньої багатоклітинності»? Однією з особливостей є взаємна залежність. Більшість клітин Анабени, ціанобактерії, мають хлорофіл і фотосинтезуються, щоб харчуватися. Анабена також виробляє клітини, звані гетероцистами, які не мають хлорофілу і не в змозі фотосинтезувати. Вони «годуються» сусідніми, «регулярними» клітинами. Гетероцисти є важливими, оскільки вони можуть приймати газ азоту, форму азоту, який «регулярні» клітини не можуть використовувати, і перетворювати його у форму, яку можуть використовувати звичайні клітини (докладніше в розділі 22). Значить, гетероцисти і регулярні клітини залежать один від одного. Принаймні деякі працівники вважають залежність одного типу клітин від іншого критерієм, який робить Anabena багатоклітинною. Ще однією особливістю, яку демонструє Анабена і яка є важливою для поняття багатоклітинності, є рух матеріалу (їжа, сполуки азоту) між клітинами організму. Це дійсно форма міжклітинного зв'язку, і багато працівників вважають міжклітинний зв'язок ключовою вимогою, що визначає багатоклітинність. До особливостей, що підсилюють зв'язок, відносяться трансклітинні зв'язки, плазмодезмати у випадку з рослинами.
Іншим критерієм, який був застосований для визначення «справжньої багатоклітинності», є тривимірна клітинна організація, тобто не просто клітини, додані в одному вимірі (нитка) або додані в двох вимірах (аркуш), або додані в трьох вимірах без визначеної програми розвитку. Справжня багатоклітинність для деяких працівників вимагає програми розвитку, яка виробляє виразну тривимірну форму, яка вимагає, щоб на розмір, форму та діяльність клітин впливало поєднання розташування (де вони знаходяться в організмі) та звідки вони прийшли (клітинна спадщина). Така структура вимагає великого спілкування і координації між осередками. Зверніть увагу, він, однак, не обов'язково вимагає тканин, що складаються з різних типів клітин.
Працівники, які вивчають еволюцію життя, вважають, що проста багатоклітинність (колонії) з'являлася не менше двадцяти п'яти разів. Складна багатоклітинність (або те, що деякі назвали б «справжньою» багатоклітинністю), але визначена без вимоги декількох типів клітин і тканин, еволюціонувала щонайменше десять разів: один раз у групі, яка призвела до тварин, одного разу в групі зелених водоростей, яка призвела до рослин, колись в іншій групі зелених водоростей, яка зробила не ведуть до рослин, двічі в червоних водоростях, двічі в бурих водоростях, три рази в грибах і кілька разів у прокаріотів. Якщо для визначення багатоклітинності потрібні тканини з декількома типами клітин (не пов'язаних з підлогою і розмноженням), то кількість багатоклітинних груп значно скорочується: просто тварини, рослини і дуже мало грибів.
Клітинна структура окремих груп неживих організмів, організмів, які можна вважати багатоклітинними, описано нижче. Виключені судинні рослини; вони розглянуті в наступному розділі.
Клітинна структура у грибів
Хоча багато біологів характеризують грибкову групу як «багатоклітинну», багато хто явно не є. Три групи грибів є коеноцитарними, і жодна з них легко характеризується як багатоклітинна. І одноклітинні гриби, які існують в декількох грибкових групах, вже точно не вважатимуться багатоклітинними. Гриби, які можуть розглядатися як володіють складною багатоклітинністю, роблять це лише для невеликої частини свого існування і лише в невеликій частині своєї структури. Майже всі гриби виробляють нитчасту структуру, гіфу (множинні гіфи), яка навряд чи багатоклітинна і краще описується як колоніальний, нитчастий організм, який не має жодної спеціалізації клітин. У трьох грибкових групах гіфи не мають поперечних стінок і є коеноцитарними. Гіфи ростуть з верхівки і можуть розгалужуватися надвоє. Розгалуження також іноді може відбуватися далеко від кінчика. Гіфи також можуть зливатися один з одним, тим самим утворюючи анастомозуючу структуру. Сформована таким чином мережа гіф називається міцелієм. Типовий міцелій - це структура живлення з динамічною (зростаючою та вмираючою одночасно), дифузною формою, яка добре підходить для отримання поживних речовин: вона має як велику площу поверхні для поглинання, так і проникає у великий об'єм навколишнього середовища, дозволяючи йому «видобувати» субстрат для поживні речовини. Харчовий міцелій є травною системою грибів і одночасно служить: (1) структурою для перетравлення їжі, (2) тонким кишечником для поглинання поживних речовин (3) кровоносною системою для переміщення поживних речовин до інших частин гриба і, зрештою, до репродуктивних структур. Зверніть увагу, що жодна з цих функцій не пов'язана з конкретними типами клітин, тканинами або органами.

Дві групи грибів вважаються клітинними, оскільки вони мають поперечні стінки. Вони називаються перегородками (сингулярна перегородка), і вони лише частково закривають клітини, оскільки вони перфоровані порами, достатньо великими, щоб дозволити багатьом матеріалам (рибосомам, мітохондріям і іноді ядрам) переміщатися з однієї клітини в іншу. Таким чином, склад цих грибів знаходиться між ценоцитарними і клітинними. Клітинна будова конкретних груп грибів описано нижче.
Хітриди
Грибкова група, що вважається найбільш примітивною (ймовірно, краще описується як група, яка розходилася в найраніший час від інших грибів) - це Хітридіомікоти (хітриди). Хітриди - це дрібні водні організми, як морські, так і прісноводні, з деякими, що існують у плівці води, що оточують частинки грунту. Більшість мікроскопічні, зазвичай розміром з типовий одноклітинний організм (менше 100 мкм). Вони можуть бути по-справжньому одноклітинними (одне ядро) або коеноцитарними (множинні ядра). Багато виробляють форми, які мають приблизно сферичну форму. Кілька хітрид утворюють так звані ризоїди, коренеподібні розширення від основного тіла, які прикріплюють їх до джерела живлення, часто пилку, спор інших організмів або однієї клітини живого або мертвого багатоклітинного організму. Крім прикріплення, ризоїди також служать для збільшення площі поверхні і дозволяють більш ефективно травлення та всмоктування. Кілька хітрид утворюють нитки (гіфи), які зрідка розгалужуються, утворюючи дуже маленький міцелій. Група відрізняється від всіх інших грибів тим, що мають джгутикові, рухливі, суперечки.
Коеноцитарні, нитчасті гриби
Дві додаткові групи грибів, хлібні цвілі (Zygomycota) і Glomeromycota, значно більші за хітриди і виробляють коеноцитарні гіфи і утворюють міцелію. Хоча обидві ці групи не мають певних типів клітин, деякі утворюють специфічні структури, які можна назвати органами, за винятком того, що вони є частинами клітин, а не складаються з клітин. Для хлібних форм ці структури, столони, ризоїди і спорангіофори, відносно великі і їх можна побачити неозброєним оком. У гломеромікоті «органи» - це набагато менші структури, але знову ж таки є частинами клітин, а не клітинами. The Glomeromycota - це гриби, які разом з рослинами утворюють ендомікоризи, асоціації між корінням рослин і грибами. При ендомікоризі гриб проникає в окремі кореневі клітини («ендо» означає всередині) і всередині цих клітин гриб утворює дві структури, деревоподібні арбускули і кулясті бульбашки (ендомікоризні гриби іноді називають везикулярно-арбускулярної мікоризою або просто VA мікоризою). Ці структури є розширеннями від ценоцитарних гіф. Арбускули - це структури, що мають деяку схожість з ризоїдами хітрид. Вони не виконують анкерної ролі, але мають значну роль у збільшенні площі контакту між грибком і рослиною, тим самим дозволяючи більшому переміщенню матеріалу між ними. Везикули - це сферичні тіла, які, як вважають, зберігають матеріали для грибка і які можуть перерости в спори.
Септат грибів - клуб і мішок грибів
Дві найбільші групи грибів, клубні гриби (Basidiomycota) та мішкові гриби (Ascomycota), не є коеноцитарними. Вони виробляють гіфи, які є перегородками, з поперечними стінками для окреслення окремих клітин. У міру розширення зростаючої верхівки гіфи ядерні відділи (мітоз) узгоджуються з клітинними відділами (цитокінезом), які виробляють нові поперечні стінки зі значною порою. Ці поперечні стінки орієнтовані перпендикулярно довгій осі гіфи. В результаті нові клітини, кожна з яких має ядро, послідовно виробляються на кінчику в міру його зростання. Кінцева клітина також здатна періодично ділитися таким чином, щоб викликати роздвоєння верхівки, розщеплюватися надвоє, утворюючи гілку. Розгалуження іноді також відбувається далеко від верхівки, в результаті наросту з раніше сформованої клітини.
За певних умов гіфи Ascomycota і Basidiomycota також можуть утворювати набагато більш компактні міцелії, зазвичай пов'язані з розмноженням, і деякі з яких наближаються у формі структур, більш характерних для багатоклітинних організмів. Ці структури можуть варіюватися в організації від аморфної маси компактних гіф (він же щільний міцелій), що називається стромою, в якій утворюються «плодові тіла» (структури, що виробляють спори) (див. Хвороба смоловидних плям), до набагато більш визначеної та детермінантної структури, такої як типовий гриб зі стеблом і ковпачок. Зазвичай спори виробляються з структури, яка якось піднята, що дозволяє посилити розгін спор вітром. Більшість із цих спороутворюючих структур називаються грибами, якщо їх видно неозброєним оком; і більшість - клубні гриби (Basidiomycota), але деякі з них - чашкові гриби (Ascomycota). У межах двох груп є найрізноманітніші форми плодових тіл, які розвиваються в результаті закономірності росту множинних гіф. Хоча ці структури чітко організовані і виробляють послідовну (детермінантну) тривимірну кінцеву форму, вони, як правило, показують мало доказів тканин або спеціалізованих типів клітин, за винятком тих, які безпосередньо пов'язані з статевим розмноженням.
Ще одна щільна компактна гіфальна структура, що виробляється перегородковими грибами, називається грибковим канатиком або ризоморфом. Він складається з декількох гіф, що йдуть паралельно і склеєні один з одним, утворюючи нитку, яка може бути кілька міліметрів в діаметрі (див. до грибкової гіфи, яка, як правило, в десять разів менше в діаметрі, ніж людський волосся). Ризоморфи є результатом повторного гіфального розгалуження під дуже низькими кутами до орієнтації батьківських гіф, отже, дозволяючи гілці залишатися тісно пов'язаною та злитою з гіфою, яка її виробляла. Результатом є структура з невеликою здатністю харчуватися (або втрачати воду) порівняно з дифузним міцелієм через зменшену площу поверхні та зменшення проникнення в навколишнє середовище. Але ризоморф спеціалізується на мобільності і дозволяє грибу проходити простір з низьким вмістом поживних речовин або екологічно вороже (наприклад, сухе) простір і потенційно прибути в більш сприятливе місце. Коли і якщо ризоморф потрапляє в зону, багату поживними речовинами, форма росту повертається до дифузного міцелію, знову придатного для засвоєння поживних речовин. Розширення ризоморфів вимагає транспортування поживних речовин для того, щоб живити кінчики гіф (де відбувається зростання), використовуючи поживні речовини, які набуваються на деякій відстані від кінчика. Цю транспортну функцію виконують спеціалізовані гіфи великого діаметру, аналогічні трубкам, виявленим у бурих водоростей і судинних рослин. Інший спеціалізований тип клітин, виявлений у ризоморфів і розташований назовні від них, - це товстостінні «обшивальні гіфи», які мають структуру, знову подібну до деяких клітин (волокон), знайдених у судинних рослин. Вони надають ризоморфу структурну цілісність і роблять його менш імовірним, що має набагато більше наслідків для ризоморфа, ніж для окремої гіфи.
Кілька грибів виробляють структури, звані склеротією, знову щільною масою гіф, але склеротії часто розвиваються в різні форми і іноді виявляють спеціалізацію між твердою зовнішньою «шкіркою» та внутрішніми клітинами. Клітини всередині часто мають значні запаси їжі, а склеротії, як правило, перезимують структури, структури, які можуть стати сплячими. Зберігається корм дозволяє склероцію, який іноді розганяється, пережити зиму і відновити зростання при поверненні сприятливих умов.
Клітинна структура водяних форм
Водяні форми, або Oomycota, схожі за формою на хлібні форми (Zygomycota), причому обидві групи складаються з коеноцитарних ниток, гіф, які в сукупності утворюють міцелій, але не показують клітинної спеціалізації, крім клітин, пов'язаних з розмноженням. До групи належать деякі важливі патогенні мікроорганізми рослин, включаючи фітофтороз картоплі, синдром раптової дубової смерті та «демпфуючі» хвороби. Незважаючи на назву, вони не цвілі (гриби), і багато з них наземні не водні (хоча представник, ймовірно, найбільш часто зустрічається, принаймні, якщо у вас є акваріум, - це білий пух, що трапляється на мертвих рибах). Водні цвілі колись були згруповані з грибами через морфологію, але зараз вони вважаються в угрупованні гетероконт, яка також включає бурі водорості та діатомові водорості.
Клітинна структура стільникових лизових форм
Під час існування одноклітинної частини клітинної слизової цвілі, очевидно, немає клітинної диференціації, але коли відбувається агрегація, клітини набувають різних долі, і можна вважати це відображенням складної багатоклітинності. Зокрема, і еволюційне значення лише деякі клітини сукупності в кінцевому підсумку виробляють репродуктивні клітини. Інші клітини, наприклад, клітини в стеблі структури, що виробляє спор, сприяють репродуктивному успіху, піднімаючи структуру вироблення спор, але не отримують еволюційної переваги для «виконання хорошої роботи», оскільки вони не обов'язково представлені в наступному поколінні. Це на відміну від більш нормального виробництва багатоклітинності, коли всі клітини багатоклітинного організму є похідними (і, отже, поділяють генетичний склад) з оригінальною «стартовою клітиною», як правило, зиготою або спорою. Через їх відносно незвичний шлях до багатоклітинності детермінатори долі клітин у клітинних слизових цвілі є областю активного вивчення. Явних візуальних/структурних відмінностей між клітинами слимака або структурою плодоношення немає, але їх положення різні, деякі зовні, деякі всередині; деякі знаходяться поруч спереду, деякі ззаду. І позиція, як видається, має важливе значення при визначенні долі клітини, включаючи позицію на момент агрегації. Більше того, є вагомі докази того, що доля клітин і «знання» положення є результатом спілкування за допомогою хімічних сигналів, одним із критеріїв, які працівники вважають визначальним аспектом складної багатоклітинності.
Клітинна структура водоростей
Водорості - це штучна угруповання водних фотосинтетичних еукаріотичних організмів, класифікація яких залишається в деякому потоці. Багато водоростей є одноклітинними або колоніальними («проста багатоклітинність») і не виявляють жодної клітинної спеціалізації, крім клітин, пов'язаних із статтю та розмноженням. Більшість цих організмів дрібні, мікроскопічні або ледь помітні неозброєним оком. Але є три групи водоростей, барвисто названих червоними водоростями, бурими водоростями та зеленими водоростями, які включають форми, які описуються як «макроводорості». Вони великі (тобто не мікроскопічні, як правило, завдовжки не менше 0,1 м) і з організованою структурою, часто складається з органів: циліндричних стебел і гілок, сплющених лопатей, повітряних мішечків, коренеподібних холдфастів і інших особливостей. Майже всі бурі водорості і більшість червоних водоростей можна було описати таким чином. Ця форма пропорційно рідше зустрічається у зелених водоростях, де багато членів менше і де значна кількість форм є одноклітинними. Використовуючи більш м'які визначення багатоклітинності, більшість макроводоростей будуть вважатися багатоклітинними. Використання більш обмежувальних визначень, зокрема вимагає міжклітинних зв'язків та зв'язку, що дозволяє взаємодіяти між клітинами, значно зменшує кількість макроводоростей, які вважаються «справді багатоклітинними». Тим не менш, виявляється, що жорстко визначена складна багатоклітинність еволюціонувала незалежно принаймні два рази в кожній з трьох груп макроводоростей (разом із трьома разів у грибів, один раз у тварин і двічі у бактерій).
Бурі водорості
Ця група раніше вважалася філумом, але тепер розглядається як сутність далі за таксономічною шкалою (клас, або сім'я, або рівень між ними). Хоча майже всі бурі водорості є макроводоростями, багатьом не вистачає клітинної диференціації та очевидної організації тканин. Структура в основному нитчаста, з нитками розгалужуються і взаємодіють один з одним, утворюючи виразну структуру, подібну до того, як певні гриби виробляють гриби з гіф. Схоже, що «справжня» багатоклітинність, з наявністю клітинної диференціації та тканин, розвивалася двічі в цій групі, один раз у лінію, яка включає камвід (фукус) і один раз у лінію, яка включає гігантську ламінарію (ламінарію). В обох цих групах є різні органи (holdfast, stipe, лопатка, повітряні бульбашки) поряд з деякою клітинною та тканинною диференціацією; зокрема, є клітини, спеціалізовані для забезпечення руху вуглеводів на великі відстані по всьому організму. Ці клітини аналогічні (не гомологічні!) з транспортними клітинами судинних рослин і мають схожі риси: вони являють собою подовжені клітини з великими діаметрами і з множинними трансклітинними зв'язками між сусідніми клітинами. Інший виразний тип клітин, також аналогічний тому, що міститься в судинних рослинам, - це клітини з потовщеними клітинними стінками, які забезпечують структурну цілісність, дозволяючи великому організму триматися разом, незважаючи на те, що його штовхають і витягують океанські течії та/або хвилі.
Червоні водорості
Група червоних водоростей має приблизно в чотири рази більше видів, ніж бурі водорості, і більш різноманітна з точки зору морфології. Значну кількість складають макроводорості з певною організованою структурою. Як це було у грибів та деяких бурих водоростей, їх тривимірна складність є наслідком зростання та взаємодії множинних ниток. Тканини можуть бути описані в деяких формах, причому циліндричні частини («стебла») мають чіткі зовнішні та внутрішні шари і мають клітини, що відрізняються розміром і пігментацією. Деякі червоні водорості утворюють кору ('crustose') і існують як покриття на поверхні різних субстратів. У багатьох коркових формах існує шарувата організація, при цьому нитки поруч з субстратом мають різні клітини від тих, що знаходяться вище. Деякі з коркових червоних водоростей розвивають шар ділильних клітин біля верхньої поверхні, дія яких дозволяє кірці потовщуватися, а також дозволяє розвивати репродуктивні структури.
Зелені водорості
З трьох водорослевих груп, що включають макроводорості, група зелених водоростей на сьогоднішній день є найбільшою (15 000 видів), з багатьма одноклітинними, простими нитчастими та колоніальними формами. Отже, макроводорості становлять набагато меншу частину від загального виду. Деякі з макроводоростей є сифоноподібними, часто складаються з багатоядерних ниток, які взаємодіють, утворюючи тривимірну форму. Більшість сучасних (кладистичних) філогеній ділять зелені водорості на дві групи, одна з яких, крім включення (деяких) зелених водоростей, містить всі організми, що розглядаються рослинами. Обидві групи демонструють різноманітні форми, включаючи деякі макроводорості з «складною» багатоклітинністю, демонструючи тривимірний ріст, клітинні зв'язки та різні типи клітин.