3.4: Шляхи C
- Page ID
- 6618

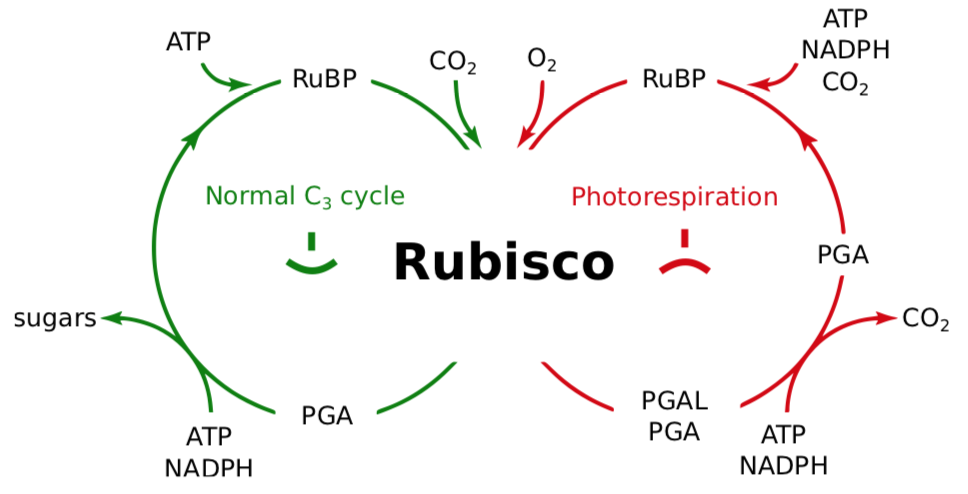
Рубіско є ферментом надзвичайно важливого значення, оскільки він починає засвоєння вуглекислого газу. На жаль, Rubisco є «дволиким», оскільки він також каталізує фотодихання (рис.\(\PageIndex{1}\)). Фотодихання означає, що рослини приймають кисень замість вуглекислого газу. Рубіско каталізує фотодихання, якщо є висока концентрація кисню (що зазвичай є результатом інтенсивної світлової стадії). Rubisco оксигенатує С\(_5\) (RubP), який перетворюється в PGA і PGAL, стаючи гліколятом. Цей гліколат повертається в цикл Кальвіна, коли клітина використовує пероксисоми і мітохондрії, і витрачає АТФ. Процес фотодихання відходить С\(_5\) і АТФ, які можуть бути більш корисними рослині іншими способами.
Якщо концентрація СО\(_2\) досить висока, асиміляція подолає фотодихання. Отже, щоб мінімізувати кількість фотодихання і зберегти їх С\(_5\) і АТФ, заводи використовують принцип Ле Шательє («Закон рівноваги») і збільшують концентрацію вуглекислого газу. Вони роблять це, тимчасово зв'язуючи вуглекислий газ з ПЕП (С\(_3\)) за допомогою ферменту карбоксилази; це призводить до\(_4\) молекул С, різних органічних кислот (таких як малат, яблучна кислота) з чотирма вуглецями в скелеті. Коли рослина потребує цього, що С\(_4\) розщеплюється на піруват (С\(_3\)) плюс вуглекислий газ, і виділення цього вуглекислого газу збільшить його концентрацію. На завершальному етапі піруват плюс АТФ реагують на відновлення ПЕП; відновлення ПЕП коштує АТФ. Весь цей процес називається «\(_\mathbf{4}\)Шляхом С» (рис.\(\PageIndex{2}\)).
Рослини, які використовують\(_4\) шлях С, відпрацьовують АТФ у своїх зусиллах відновити ПЕП, але вони все ще перевершують\(_3\) фотореспіраторні С-рослини, коли спостерігається інтенсивне світло та/або висока температура і, отже, висока концентрація кисню. Саме тому в тропічному кліматі кращими є С\(_4\) -культури.

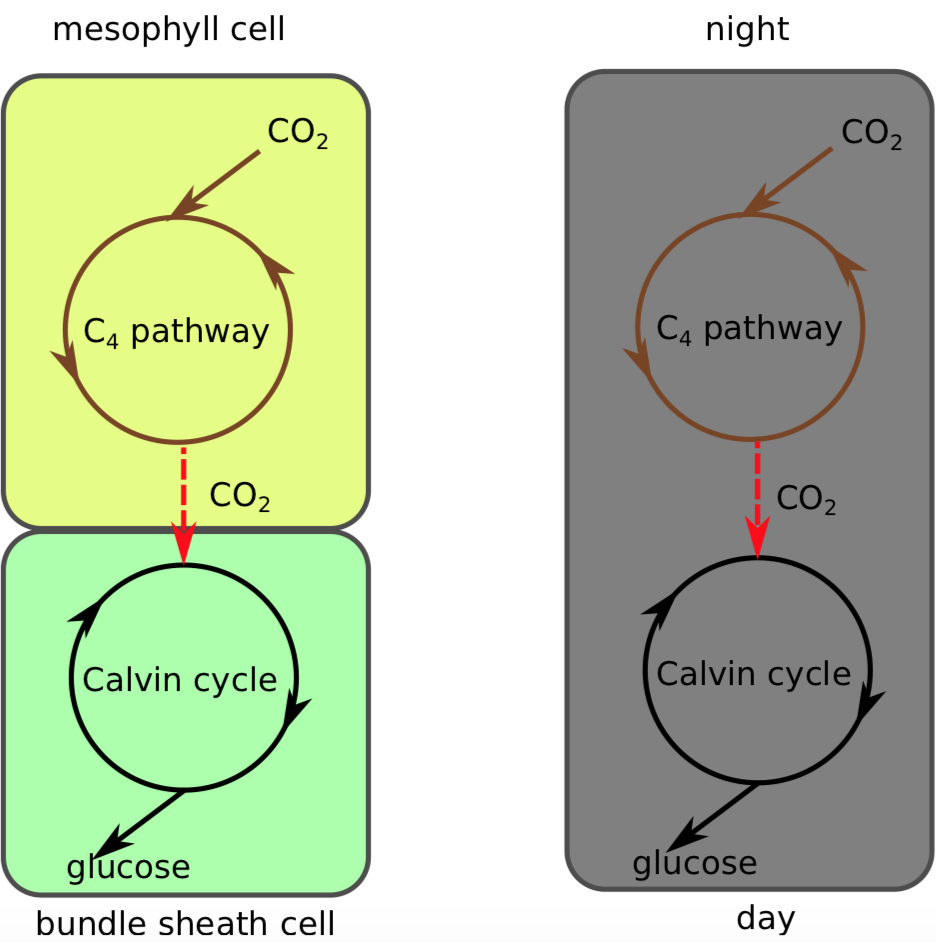
Дві групи рослин використовують\(_4\) шлях С. Багато пустельних або сухих рослин - це CAM-рослини, які вночі проїжджають\(_4\) шлях C. Вони роблять тимчасовий поділ між накопиченням вуглекислого газу і фотосинтезом. Кулачкові рослини складають сім відсотків рослинного різноманіття, і мають 17 000 різних видів (наприклад, ананас (ананас), кактуси, кактуси; рослина нефрит, толстянка і їх родичі).
«Класичні»\(_4\) рослини С проганяють\(_4\) шлях С в клітині мезофілу листя, тоді як їх С\(_3\) знаходиться в так званих клітинках оболонки пучка. Це просторове, а не тимчасове поділ. Ці\(_4\) С-рослини складають три відсотки біорізноманіття рослин і мають понад 7000 різних видів (наприклад, кукурудза, Зея; сорго, сорго та їх родичі). В цілому обидва варіанти\(_4\) шляху С пов'язані з концентрацією вуглекислого газу, просторової або тимчасової (рис.\(\PageIndex{3}\)). Обидва вони називаються «вуглецево-концентрованими механізмами», або ССМ.
Є рослини, які здатні керувати\(_4\) шляхами C\(_3\) і C (наприклад, дерево автографа, Clusia), і рослини, що мають як «класичні» варіанти C, так\(_4\) і CAM (наприклад, Portulacaria).