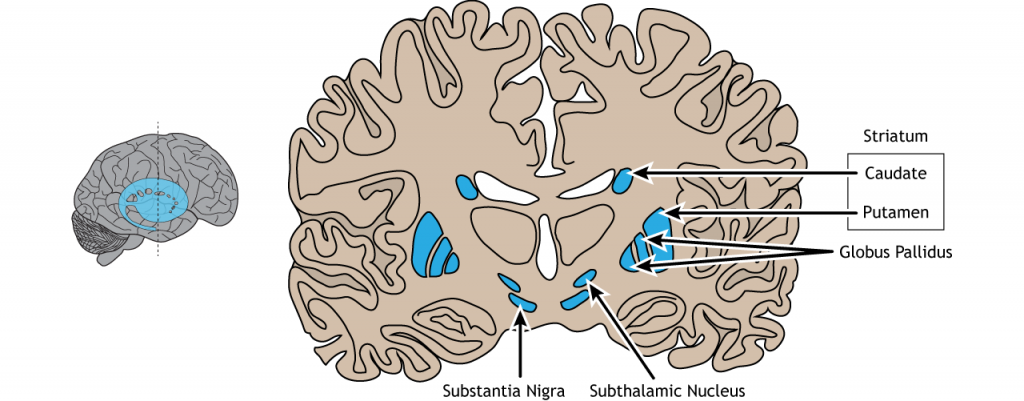
Базальні ганглії - це група підкіркових ядер, що означає групи нейронів, які лежать нижче кори головного мозку. Базальні ганглії складаються з стриатуму, який складається з хвостатого ядра та путамена, глобуса паллідуса, субталамічного ядра та чорної субстанції. Базальні ганглії в першу чергу пов'язані з руховим контролем, оскільки рухові розлади, такі як хвороби Паркінсона або Хантінгтона, походять від дисфункція нейронів всередині базальних гангліїв. Для добровільної рухової поведінки базальні ганглії беруть участь у ініціації або придушенні поведінки і можуть регулювати рух за допомогою модулюючої активності в таламусі та корі. Окрім рухового контролю, базальні ганглії також спілкуються з немоторними областями кори головного мозку та відіграють певну роль в інших поведінках, таких як емоційна та когнітивна обробка.
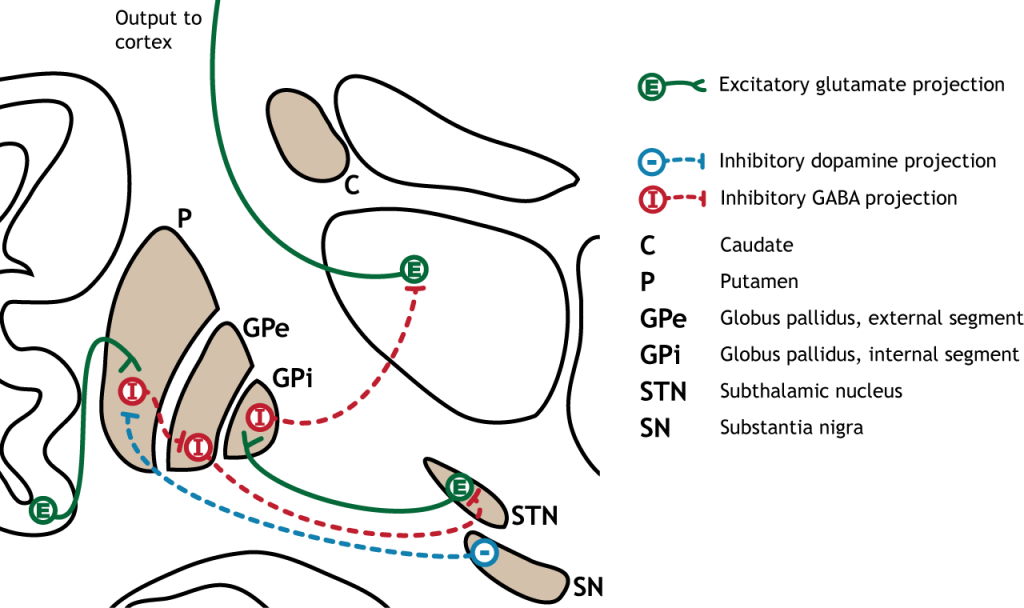
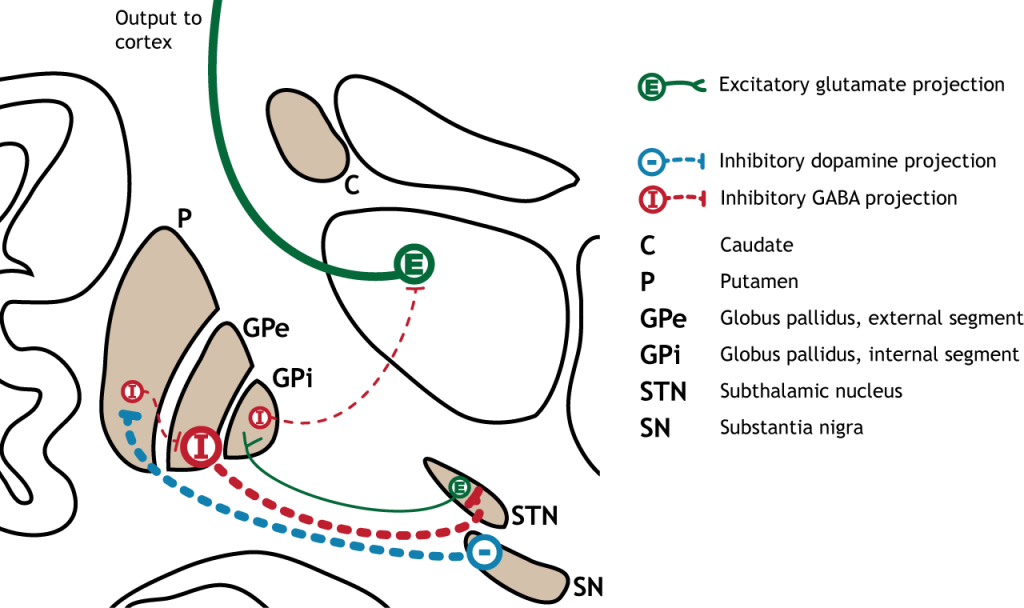
Малюнок 27.1. Базальні ганглії - це підкіркові структури, розташовані біля основи переднього мозку. Вони складаються з хвостатого і путамена, які обидва складають стріатум, а також глобус паллідус, чорна субстанція та субталамічне ядро. «Базальна ганглія' Кейсі Хенлі ліцензовано на умовах Ліцензії Creative Commons Зазначення Авторства Некомерційна Частка На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.
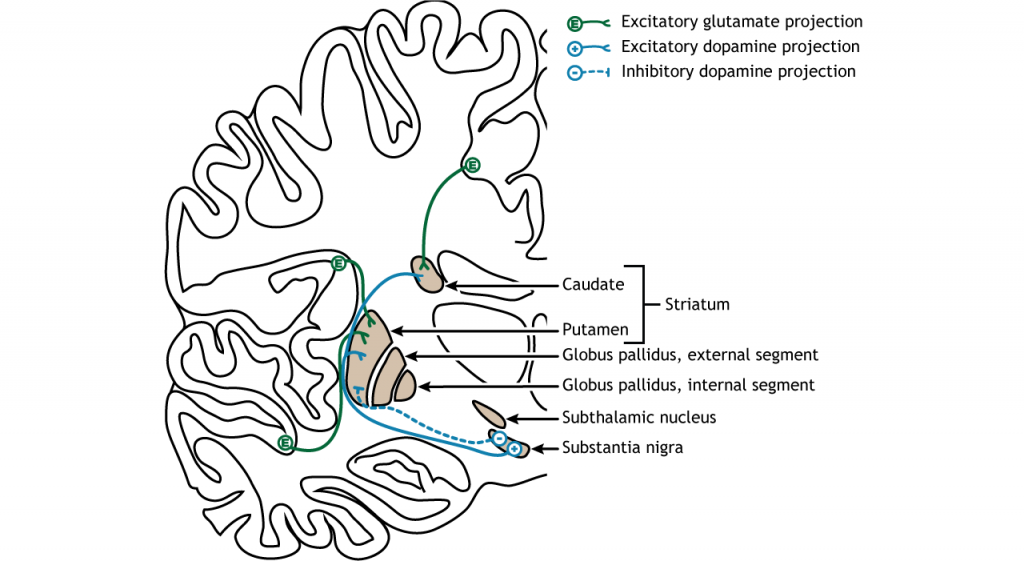
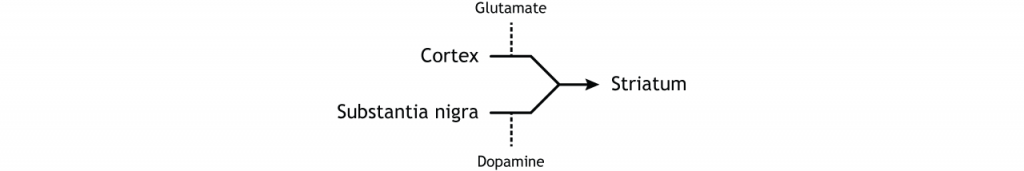
Більшість інформації, що обробляється базальними гангліями, надходить через смугастий. Основне джерело введення в базальні ганглії - з кори головного мозку. Цей вхід є глуматергічним і, отже, збудливим. Нігра субстанції також є областю з критичними проекціями до смугастої смуги і є основним джерелом дофамінергічного введення. Дофамін відіграє важливу роль у функції базальних гангліїв. Хвороба Паркінсона призводить, коли нейрони дофаміну в чорній субстанції вироджуються і більше не надсилають відповідні входи в смугу. Проекції дофаміну можуть мати або збудливу, або інгібуючу дію в смузі, залежно від типу метаботропного дофамінового рецептора, який висловлює стриатальний нейрон. Дія дофаміну на нейрон, який виражає рецептор D1, є збудливим. Дія дофаміну на нейрон, який виражає рецептор D2, є інгібуючим.
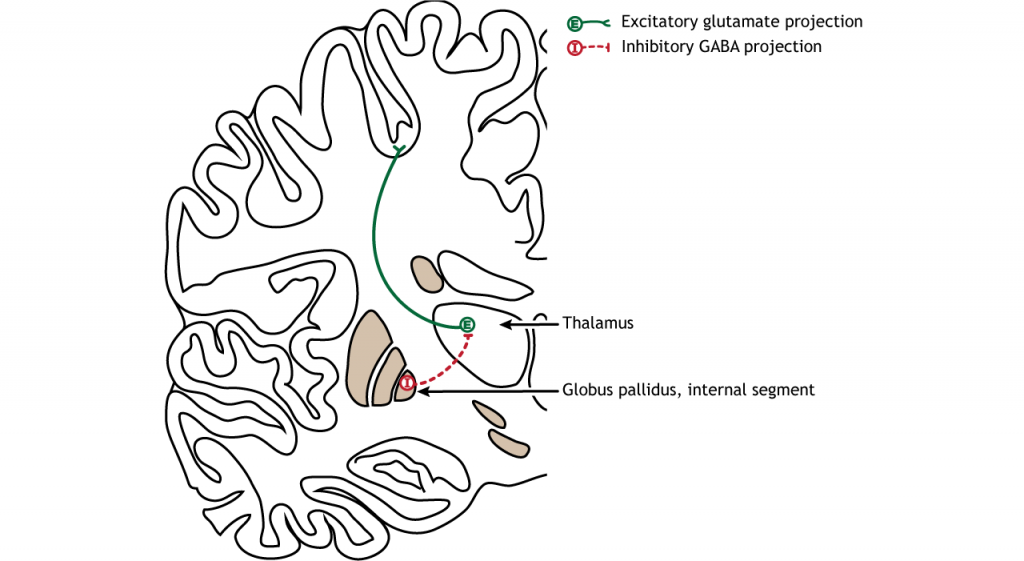
Первинною вихідною областю базальних гангліїв є внутрішній сегмент globus pallidus. Ця область посилає гальмівні GABaERgic проекції до ядер в таламусі. Цей гальмівний вихід має тонічну, постійну швидкість стрільби, що дозволяє базальним гангліям вихід як збільшуватися, так і зменшуватися залежно від ситуації. Потім таламус проектує назад до кори головного мозку, насамперед до рухових областей.
Існує кілька зв'язків у базальних структурах гангліїв, а також. Для управління двигуном існують дві основні схеми: прямий шлях і непрямий шлях. Ці схеми мають протилежні дії при активації коркових нейронів. Схеми також модулюються вивільненням дофаміну чорною субстанцією в смугу. Вважається, що різні механізми управління дозволяють точно налаштований баланс між прямим і непрямим ланцюгами, що дозволяє досконалити контроль руху.
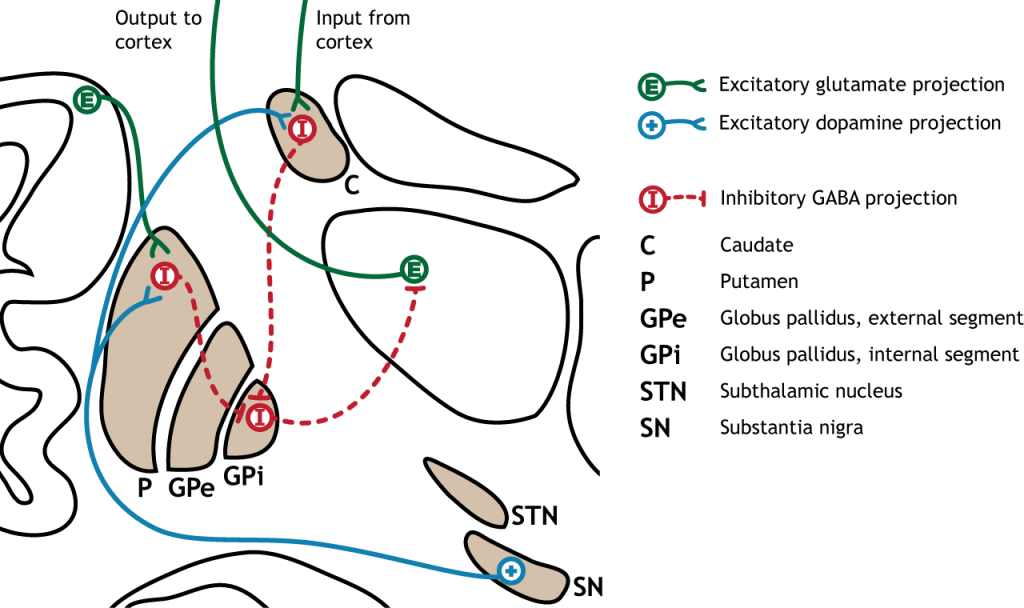
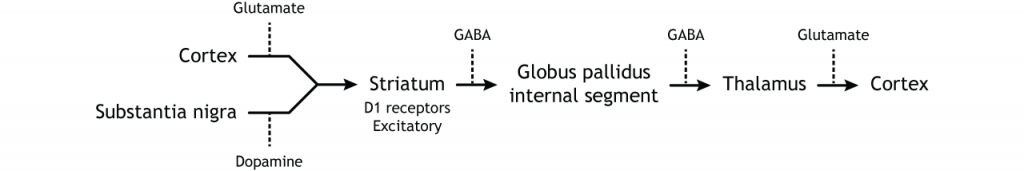
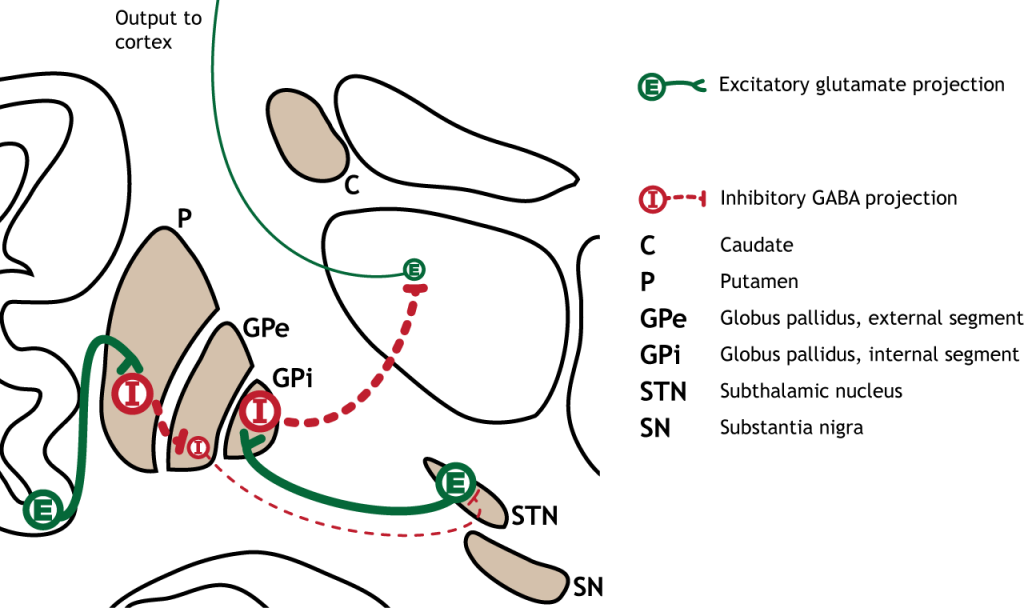
Прямий шлях починається в стріатумі, який посилає гальмівні проекції на внутрішній сегмент глобуса паллідуса (GPi). Потім GPi посилає інгібуючий вихід в таламус.
Малюнок 27.6. Прямий шлях у базальних гангліях складається з збудливого введення з кори через глутаматну дію або субстанцію nigra через дію дофаміну, яка синапсує на інгібуючі нейрони в смугасті. Стриатальні нейрони проектують на внутрішній сегмент глобуса pallidus (GPi). Потім GPi посилає інгібуючий вихід в таламус. «Прямий шлях до базальних гангліїв» Кейсі Хенлі ліцензовано на умовах Ліцензії Creative Commons Із Зазначенням Авторства Некомерційна Частка На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.Малюнок 27.7. Кора посилає глутаматні проекції до смугастої оболонки. Нігра субстанції посилає дофамінові проекції до смуги, які є збудливими, впливаючи на рецептори D1 в нейрони, що беруть участь у прямому шляху. Стріатум посилає проекції ГАМК на внутрішній сегмент глобуса pallidus (GPi). ГПі посилає ГАМК проекції на таламус. Таламус посилає проекції глутамату в кору. «Прямий шлях базальних гангліїв — текст» Кейсі Хенлі ліцензовано на умовах Ліцензії Creative Commons Із Зазначенням Авторства Некомерційна Частка На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.
Активація прямого шляху
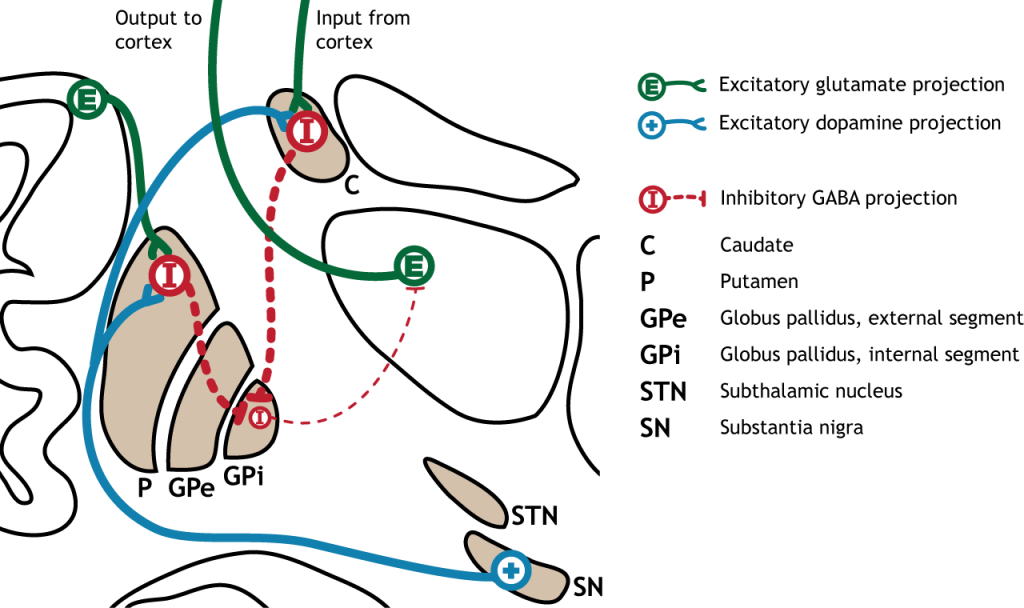
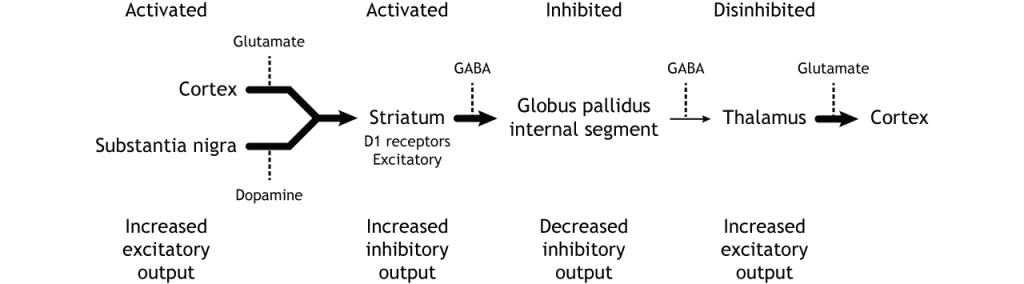
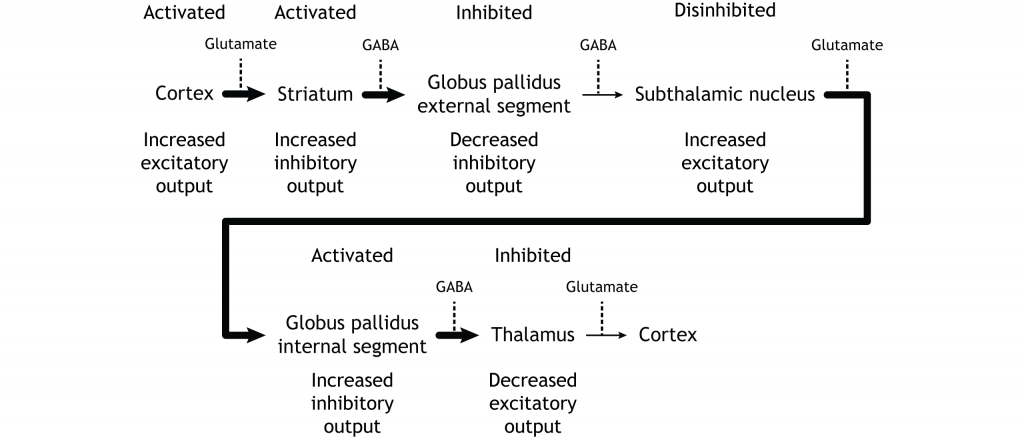
Коли введення або з кори або субстанції чорної збільшується в інтенсивності, прямий шлях активується. Нейрони в смузі, що беруть участь у прямому шляху, експресують метаботропний дофаміновий рецептор D1, і активація цього рецептора є збудливою. Тому проекції як з кори, так і від чорної субстанції активують нейрони в смугастому. Ці нейрони інгібують і вивільняють ГАМК на внутрішній сегмент глобусу pallidus (GPi). Як описано вище, нейрони в ГПі є інгібуючими, вивільняючи ГАМК на таламус. Активація смугастих нейронів пригнічує нейрони в ГПі, звільняючи гальмування на таламусі. Пригнічення інгібуючої області називається розгальмуванням. Тому активація прямого шляху призводить до збільшення виходу з таламуса, оскільки він розгальмований.
Малюнок 27.8. Активація прямого шляху або збільшеним входом з кори або субстанції, або substantia nigra призводить до збільшення інгібуючого виходу зі смугастого в GPI. Інгібування на ГПі призводить до менш гальмівного введення в таламус, викликаючи підвищений вихід з таламуса в кору. «Активація прямого шляху» Кейсі Хенлі ліцензується на умовах Ліцензії Creative Commons Із Зазначенням Авторства Некомерційна Поширення На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.Малюнок 27.9. Коли або кора, або чорна субстанція активуються, вони посилають підвищений збуджуючий вихід в смугастий, який виражає збудливі D1 рецептори в нейрони, що беруть участь у прямому шляху. Цей вхід активує стріатум, який посилає збільшені інгібуючі проекції на GPI. Пригнічений GPI посилає знижені інгібуючі проекції в таламус, знешкоджуючи таламус. Потім таламус посилає підвищений збудливий вихід в кору. «Активація прямого шляху — текст» Кейсі Хенлі ліцензується на умовах Ліцензії Creative Commons Із Зазначенням Авторства Некомерційна Частка На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.
Непрямий шлях
Непрямий шлях трохи складніше. Як і прямий шлях, вхід в базальні ганглії виникає з кори і субстанції nigra, але всередині базальних гангліїв є більше внутрішніх зв'язків, що відбувається в прямому шляху. Інгібуючі нейрони в стріатумі, що беруть участь в непрямому шляху проектуються на зовнішній сегмент глобуса pallidus (GpE). ГАМК-ергічні нейрони в GpE проектують до субталамічного ядра, яке потім посилає збудливий вихід до GPI, який виводиться в таламус.
Малюнок 27.10. Непрямий шлях у базальних гангліях складається з збудливого введення з кори за допомогою глутамату дії або інгібуючого введення від нігри субстанції через дію допаміну, яка синапсує на інгібуючі нейрони в смугастому. Стриатальні нейрони проектують на зовнішній сегмент глобуса pallidus (GpE). GpE посилає інгібуючий вихід до субталамічного ядра, яке мало збуджуючі проекції на GPi. Потім GPi посилає інгібуючий вихід в таламус. «Непрямий шлях базальних гангліїв» Кейсі Хенлі ліцензовано на умовах Ліцензії Creative Commons Із Зазначенням Авторства Некомерційна Частка На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.Малюнок 27.11. Кора посилає глутаматні проекції до смугастої оболонки. The substantia nigra посилає дофамінові проекції до смуги, які є інгібуючими, впливаючи на рецептори D2 в нейрони, що беруть участь в непрямому шляху. Стріатум посилає проекції ГАМК на зовнішній сегмент глобуса pallidus (GpE). GPE надсилає проекції ГАМК до субталамічного ядра. Субталамическое ядро посилає проекції глутамату на GPi. GPi посилають GABA-проекції на таламус. Таламус посилає проекції глутамату в кору. «Непрямий шлях базальних гангліїв — текст» Кейсі Хенлі ліцензовано на умовах Ліцензії Creative Commons Із Зазначенням Авторства Некомерційна Частка На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.
Активація непрямого шляху
Непрямий шлях активується збудливим корковим введенням, активуючи гальмуючі стриатальні нейрони. Це призводить до гальмування нейронів GpE, що призводить до розгальмування збудливих нейронів в субталамічному ядрі. Збуджуючий вихід з субталамічного ядра в ГПі збільшує пригнічення таламуса, що призводить до зниження виходу таламуса в кору.
Малюнок 27.12. Активація непрямого шляху збуджуючим корковим введенням в смугасте тіло призводить до збільшення інгібуючого виходу в GPE. Пригнічений GPE посилає знижений інгібуючий вихід до субталамічного ядра, викликаючи збільшення збуджуючого виходу з субталамічного ядра до GPi. Активація ГПі пригнічує таламус, в результаті чого знижується вихід з таламуса в кору. «Активація непрямим шляхом» Кейсі Хенлі ліцензується на умовах Ліцензії Creative Commons Зазначення Авторства Некомерційна Поширення На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.Малюнок 27.13. Коли кора активується, вона посилає підвищений збудливий вихід в смугасте тіло. Цей вхід активує стріатум, який посилає збільшені інгібуючі проекції на GPE. Пригнічений GPE посилає знижені інгібуючі проекції до субталамічного ядра, дезінгібуючи область. Потім субталамічне ядро посилає підвищений збуджуючий вихід до GPi. Активований ГПі посилає підвищені інгібуючі проекції в таламус, який посилає знижений збуджуючий вихід в кору. «Активація непрямим шляхом — текст» Кейсі Хенлі ліцензується на умовах Ліцензії Creative Commons Зазначення Авторства Некомерційна Поширення На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.
Інгібування непрямого шляху
Непрямий шлях може бути пригнічений вивільненням дофаміну з чорної субстанції. Нейрони в смузі, що беруть участь в непрямому шляху, експресують D2 метаботропний дофаміновий рецептор. Активація цього рецептора є інгібуючою. Якщо непрямий шлях гальмується проекціями дофаміну з чорної субстанції, інгібуючі стриатальні нейрони інгібуються. Це призводить до дезінгібування нейронів GpE, що призводить до гальмування збуджуючих нейронів в субталамічному ядрі. Це зниження збуджуючого виходу в ГПі зменшує гальмування таламуса, що призводить до збільшення таламуса виходу в кору.
Малюнок 27.14. Інгібування непрямого шляху шляхом інгібуючого введення з чорної субстанції до смугастої смуги призводить до зниження інгібуючого виходу в GpE. Розгальмований GPE посилає підвищений інгібуючий вихід до субталамічного ядра, викликаючи зниження збудливого виходу з субталамічного ядра до GPi. Зниження активації ГПі звільняє гальмування на таламусі, в результаті чого збільшується вихід з таламуса в кору. «Непрямий шлях гальмування» Кейсі Хенлі ліцензовано на умовах Ліцензії Creative Commons Із Зазначенням Авторства Некомерційна Частка На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.Малюнок 27.15. Коли субстанція nigra активується, вона посилає підвищений інгібуючий вихід в смугастий, який виражає інгібуючі рецептори De в нейрони, що беруть участь в непрямому шляху. Цей вхід гальмує стріатум, який посилає зменшені інгібуючі проекції до GpE. Розгальмований GPE посилає підвищені інгібуючі проекції до субталамічного ядра, пригнічуючи область. Потім субталамічне ядро посилає знижений збуджуючий вихід до GPi. Деактивований ГПі посилає зменшені інгібуючі проекції в таламус, який посилає підвищений збуджуючий вихід в кору. «Непрямий шлях гальмування — текст» Кейсі Хенлі ліцензується на умовах Ліцензії Creative Commons Зазначення Авторства Некомерційна Частка На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.
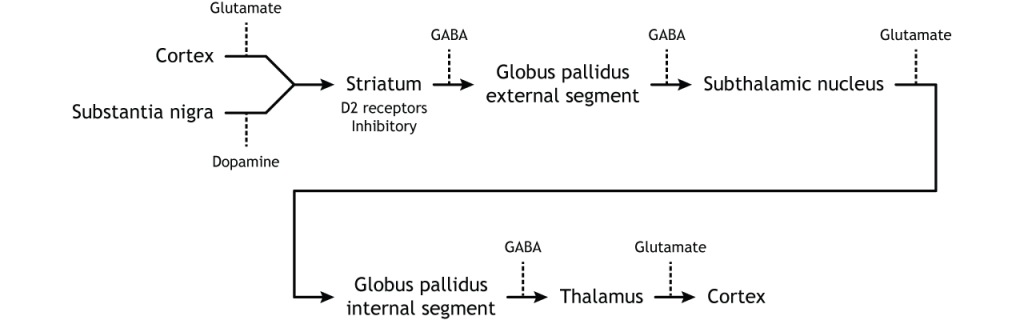
Резюме внутрішньої обробки
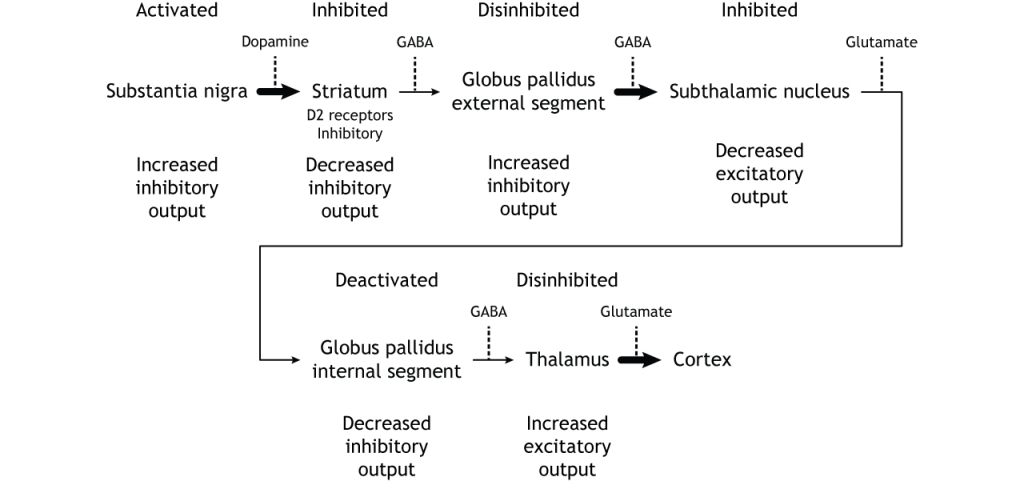
Щоб скласти все це разом, є вхід в смугастий з двох різних місць: кора (глутамат) і субстанція нігра (дофамін).
Коркова активація прямого шляху призводить до збільшення виходу таламуса
Коркова активація непрямого шляху призводить до зниження таламічного виходу
Активація чорної субстанції (через D1) прямого шляху призводить до збільшення виходу таламуса
Інгібування нігри субстанції (через D2) непрямого шляху призводить до збільшення таламічного виходу
Саме поєднання цих шляхів дозволяє точно контролювати рух двигуна.
Петлі через базальні ганглії
Існує кілька контурів, які проходять через базальні ганглії:
Схема двигуна, яка відіграє роль у добровільному русі
Окоруховий контур, який відіграє роль у русі очей
Асоціативний контур, який відіграє певну роль у виконавчих функціях, таких як гальмування поведінки (запобігання імпульсивної поведінки), планування та вирішення проблем, а також посередництво соціально відповідної поведінки
Лімбічний або емоційний контур, який відіграє певну роль у обробці емоцій та винагороди.
Хоча схеми використовують різні ланцюги всередині базальних гангліїв, загальна петля однакова: корковий вхід у смугастий призводить до внутрішньої обробки в базальних структурах гангліїв. Базальні ганглії виводять проекції від палліда до таламуса, який потім проектує назад в кору. Важливо визнати, що базальні ганглії відіграють важливу роль в ряді функцій. Наприклад, ліки, які використовуються для лікування Паркінсона, іноді можуть призвести до прояву порушень імпульсного контролю, в результаті дофамінергічних змін лімбічної петлі через базальні ганглії.
Малюнок 27.16. Петлі через базальні ганглії мають різні функції, але слідують за тією ж загальною схемою. Кора входів в стріатум. Відбувається внутрішня обробка через ланцюги базальних гангліїв, а потім вихід з палліда проектує на таламус, який відправляє вихід в кору. «Базальні гангліозні петлі» Кейсі Хенлі ліцензовано на умовах Ліцензії Creative Commons Зазначення Авторства Некомерційна Частка На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна.
Ключові виноси
Підкіркові базальні ядра гангліїв отримують інформацію з кори і відправляють вихід в таламус.
Руховий контроль через базальні ганглії відбувається як прямим, так і непрямим шляхами
Розгальмування - це коли інгібуюча область сама пригнічується
Базальні ганглії найбільш відомі своєю роллю в руховому контролі, але також мають вирішальне значення для емоцій та поведінкового гальмування.