11.2: Реплікація ДНК
- Page ID
- 3918

Цілі навчання
- Поясніть значення напівконсервативної реплікації ДНК
- Поясніть, чому реплікація ДНК є двонаправленою і включає як провідну, так і відстаючу нитку
- Поясніть, чому утворюються фрагменти Окадзакі
- Опишіть процес реплікації ДНК та функції задіяних ферментів
- Визначте відмінності між реплікацією ДНК у бактерій та еукаріотів
- Поясніть процес тиражування кочення кола
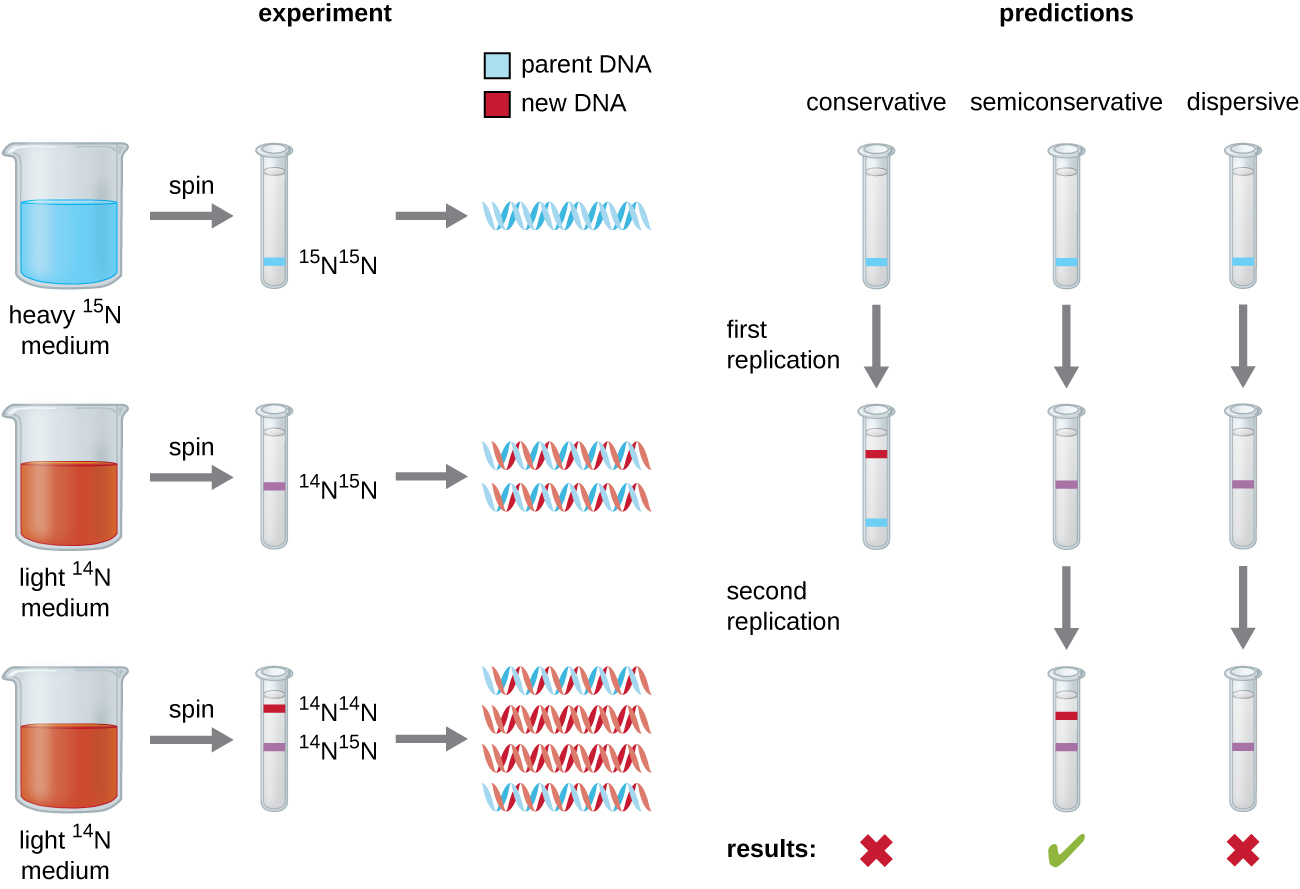
Роз'яснення структури подвійної спіралі Джеймсом Уотсоном і Френсісом Криком в 1953 році дало натяк на те, як копіюється ДНК в процесі реплікації. Відокремлення ниток подвійної спіралі надало б два шаблони для синтезу нових взаємодоповнюючих ниток, але як саме були побудовані нові молекули ДНК, все ще незрозуміло. В одній моделі, напівконсервативної реплікації, дві нитки подвійної спіралі відокремлюються під час реплікації ДНК, і кожна нитка служить шаблоном, з якого копіюється нова комплементарна нитка; після реплікації кожна двониткова ДНК включає одну батьківську або «стару» нитку і одну «нову» нитку. Також було запропоновано дві конкуруючі моделі: консервативна і дисперсійна, які показані на малюнку\(\PageIndex{1}\).

Метью Месельсон (1930—) та Франклін Шталь (1929—) розробили експеримент у 1958 році, щоб перевірити, яка з цих моделей правильно представляє реплікацію ДНК (рис.\(\PageIndex{2}\)). Вони вирощували кишкову паличку протягом декількох поколінь у середовищі, що містить «важкий» ізотоп азоту (15 N), який був включений в азотисті основи і, врешті-решт, в ДНК. Це позначено батьківською ДНК. Потім культура кишкової палички була переведена в середовище, що містить 14 N, і дозволила рости протягом одного покоління. Клітини були зібрані, а ДНК виділена. ДНК відокремлювалася ультрацентрифугуванням, в ході якої ДНК утворила смуги відповідно до її щільності. Очікується, що ДНК, вирощена в 15 N, утворює смугу в більш високому положенні щільності, ніж вирощена в 14 Н. Месельсон і Шталь зазначили, що після одного покоління зростання в 14 Н, одна смуга, що спостерігається, була проміжною в положенні між ДНК клітин, вирощених виключно в 15 N або 14 N. Це припускало або напівконсервативний, або дисперсійний режим реплікації. Деяким клітинам дозволили рости ще на одне покоління в 14 Н і знову закручувалися. ДНК, зібрана з клітин, вирощених протягом двох поколінь в 14 N, утворила дві смуги: одна група ДНК перебувала в проміжному положенні між 15 N і 14 N, а інша відповідала смузі 14 N ДНК. Ці результати можна пояснити лише в тому випадку, якщо ДНК реплікується напівконсервативним способом. Тому дві інші моделі були виключені. В результаті цього експерименту ми тепер знаємо, що під час реплікації ДНК кожна з двох ниток, що складають подвійну спіраль, служить шаблоном, з якого копіюються нові нитки. Нова пасмо буде доповнювати батьківську або «стару» пасмо. Отримані молекули ДНК мають однакову послідовність і поділяються порівну на дві дочірні клітини.
Вправа\(\PageIndex{1}\)
Яким був би висновок експерименту Месельсона і Шталя, якби після першого покоління вони знайшли дві смуги ДНК?
Реплікація ДНК у бактерій
Реплікація ДНК була добре вивчена у бактерій насамперед через невеликий розмір генома та мутантів, які доступні. E. coli має 4,6 мільйона пар основ (Mbp) в одній круговій хромосомі, і вся вона тиражується приблизно за 42 хвилини, починаючи від одного походження реплікації і протікаючи по колу двонаправлено (тобто в обох напрямках). Це означає, що в секунду додається приблизно 1000 нуклеотидів. Процес досить швидкий і відбувається з невеликою кількістю помилок.
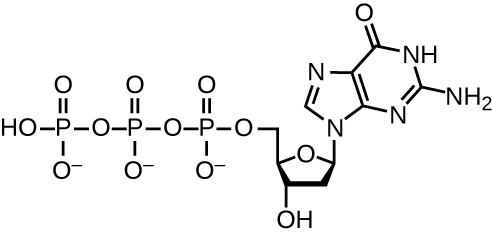
Для реплікації ДНК використовується велика кількість білків і ферментів (табл.\(\PageIndex{1}\)). Одним з ключових гравців є фермент ДНК-полімераза, також відомий як ДНК-пол. У бактерій відомі три основні типи ДНК-полімераз: ДНК пол I, ДНК пол II і ДНК пол III. Зараз відомо, що DNA pol III є ферментом, необхідним для синтезу ДНК; ДНК pol I і DNA pol II в першу чергу необхідні для відновлення. DNA pol III додає дезоксирибонуклеотиди кожен доповнює нуклеотид на нитку шаблону, один за іншим до групи 3'-OH зростаючого ланцюга ДНК. Додавання цих нуклеотидів вимагає енергії. Ця енергія присутня в зв'язках трьох фосфатних груп, прикріплених до кожного нуклеотиду (трифосфатного нуклеотиду), подібно до того, як енергія зберігається в фосфатних зв'язках аденозинтрифосфату (АТФ) (рис.\(\PageIndex{3}\)). Коли зв'язок між фосфатами порушується і дифосфат вивільняється, енергія, що виділяється, дозволяє сформувати ковалентний фосфодіефірний зв'язок шляхом синтезу зневоднення між вхідним нуклеотидом і вільною групою 3'-OH на зростаючому ланцюжку ДНК.
Ініціація
Ініціювання реплікації відбувається при певній нуклеотидній послідовності, яка називається походженням реплікації, де різні білки зв'язуються, щоб почати процес реплікації. Кишкова паличка має єдине походження реплікації (як і більшість прокаріотів), зване OriC, на своїй одній хромосомі. Походження реплікації становить приблизно 245 пар основи і багате послідовностями аденін-тиміну (АТ).
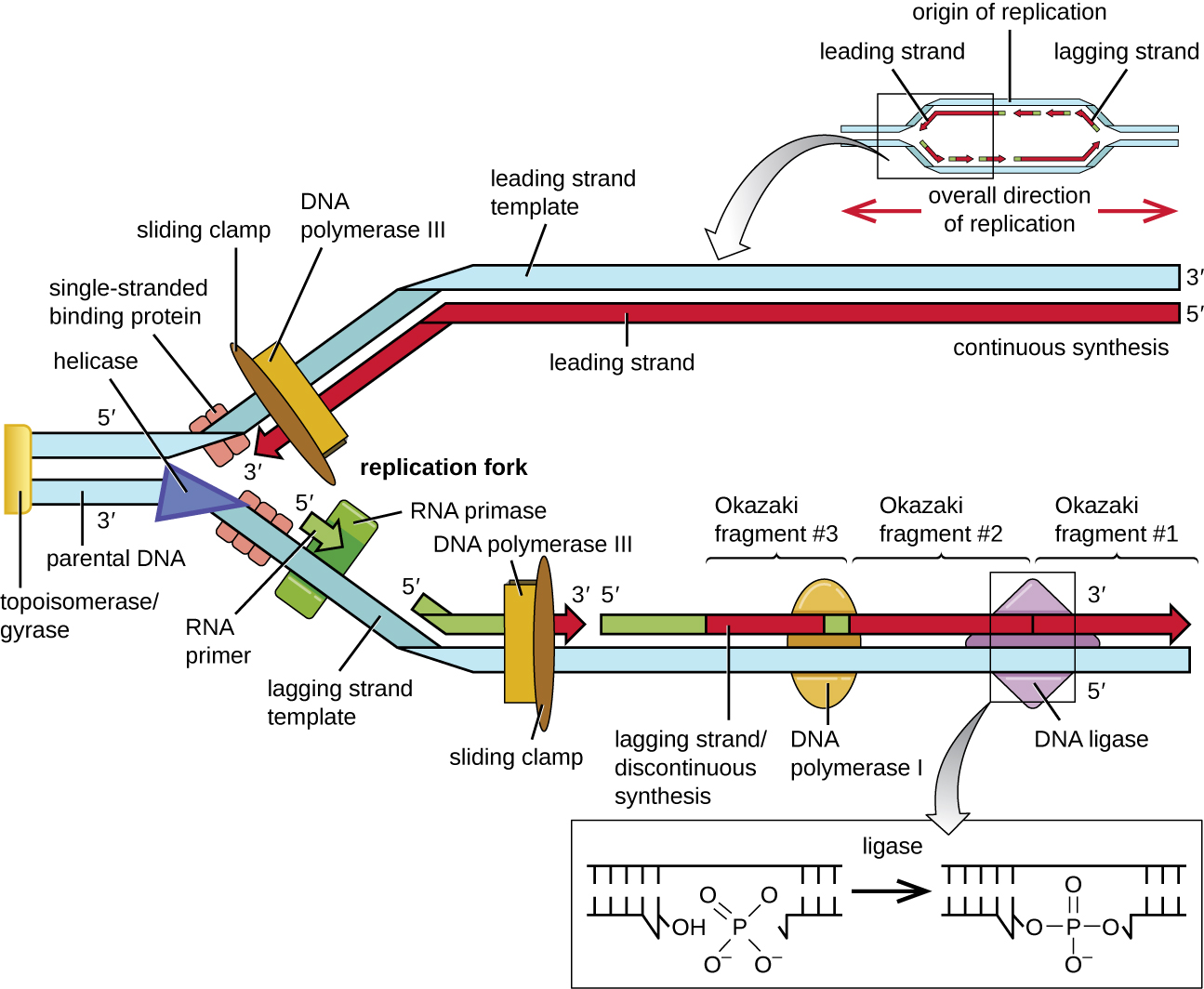
Деякі білки, які зв'язуються з походженням реплікації, важливі для того, щоб зробити одноланцюгові області ДНК доступними для реплікації. Хромосомна ДНК, як правило, обертається навколо гістонів (у еукаріотів та архей) або гістоноподібних білків (у бактерій), і перекручується, або широко загортається і скручується на себе. Ця упаковка робить інформацію в молекулі ДНК недоступною. Однак ферменти, звані топоізомеразами, змінюють форму і суперколювання хромосоми. Для початку реплікації бактеріальної ДНК суперспіральна хромосома розслабляється топоізомеразою II, яку також називають ДНК-гіразою. Потім фермент під назвою helicase відокремлює нитки ДНК, розриваючи водневі зв'язки між парами азотистих основ. Нагадаємо, що послідовності АТ мають менше водневих зв'язків і, отже, мають слабші взаємодії, ніж послідовності гуанін-цитозин (ГК). Ці ферменти вимагають гідролізу АТФ. Коли ДНК відкривається, утворюються Y-подібні структури, які називаються реплікаційною вилкою. У початку реплікації утворюються дві вилки реплікації, що дозволяє двонаправлену реплікацію та формування структури, яка виглядає як міхур при перегляді за допомогою просвічувального електронного мікроскопа; в результаті ця структура називається бульбашкою реплікації. ДНК біля кожної вилки реплікації покрита одноланцюговими зв'язуючими білками, щоб запобігти перемотуванню одноцепочечной ДНК в подвійну спіраль.
Після того, як одноцепочечная ДНК буде доступна на початку реплікації, може початися реплікація ДНК. Однак ДНК pol III здатний додавати нуклеотиди тільки в напрямку 5' до 3' (нова нитка ДНК може бути розширена тільки в цьому напрямку). Це пов'язано з тим, що ДНК-полімераза вимагає вільної групи 3'-OH, до якої вона може додавати нуклеотиди, утворюючи ковалентний фосфодіефірний зв'язок між кінцем 3'-OH та 5' фосфатом наступного нуклеотиду. Це також означає, що він не може додавати нуклеотиди, якщо вільна група 3'-OH недоступна, що стосується однієї нитки ДНК. Проблема вирішується за допомогою послідовності РНК, що забезпечує вільний кінець 3'-ОН. Оскільки ця послідовність дозволяє почати синтез ДНК, її відповідним чином називають праймером. Грунтовка має довжину від п'яти до 10 нуклеотидів і доповнює батьківську або шаблонну ДНК. Він синтезується РНК-примазою, яка є РНК-полімеразою. На відміну від ДНК-полімераз, РНК-полімерази не потребують вільної групи 3'-ОН для синтезу молекули РНК. Тепер, коли праймер забезпечує вільну групу 3'-OH, ДНК-полімераза III тепер може розширити цей праймер РНК, додаючи нуклеотиди ДНК один за іншим, які доповнюють нитку шаблону (рис.\(\PageIndex{1}\)).
Подовження
Під час подовження в реплікації ДНК приєднання нуклеотидів відбувається з максимальною швидкістю близько 1000 нуклеотидів в секунду. ДНК-полімераза III може поширюватися лише в напрямку 5' до 3', що створює проблему на розвилці реплікації. Подвійна спіраль ДНК є антипаралельною; тобто одна нитка орієнтована в напрямку 5' до 3', а інша орієнтована в напрямку 3' до 5' (див. Структура і функція ДНК). Під час реплікації одна нитка, яка доповнює батьківську ДНК 3 'до 5', синтезується безперервно до вилки реплікації, оскільки полімераза може додавати нуклеотиди в цьому напрямку. Ця безперервно синтезована пасмо відома як провідна пасмо. Інша нитка, що доповнює батьківську ДНК від 5 'до 3', відростає від вилки реплікації, тому полімераза повинна рухатися назад до вилки реплікації, щоб почати додавати основи до нового праймера, знову в напрямку від вилки реплікації. Робить це до тих пір, поки не вдариться в раніше синтезовану пасмо, а потім знову повернеться назад (рис.\(\PageIndex{4}\)). Ці кроки виробляють невеликі фрагменти послідовності ДНК, відомі як фрагменти Окадзакі, кожен розділений праймером РНК. Окадзакі фрагменти названі на честь японської дослідницької групи і подружньої пари Рейдзі і Цунеко Окадзакі, які вперше виявили їх в 1966 році. Пасмо з фрагментами Окадзакі відома як відстаюча пасмо, і її синтез, як кажуть, переривчастий.
Провідну пасмо можна продовжити з одного праймера самостійно, тоді як відстає пасмо потребує нового праймера для кожного з коротких фрагментів Окадзакі. Загальний напрямок відставання пасма буде 3' до 5', а провідної пасма 5' до 3'. Білок, який називається ковзаючим затискачем, утримує ДНК-полімеразу на місці, оскільки він продовжує додавати нуклеотиди. Зсувний затискач являє собою кільцеподібний білок, який зв'язується з ДНК і утримує полімеразу на місці. Крім своєї ролі в ініціації, топоізомераза також запобігає перемотуванню подвійної спіралі ДНК перед виделкою реплікації, коли ДНК відкривається; це робить, викликаючи тимчасові уколи в спіралі ДНК, а потім перегерметизуючи її. У міру протікання синтезу праймери РНК замінюються ДНК. Праймери видаляються екзонуклеазної активністю ДНК-полімерази I, а проміжки заповнюються. Ніки, які залишаються між новосинтезованою ДНК (яка замінила праймер РНК) та раніше синтезованою ДНК, запечатані ферментом ДНК-лігазою, яка каталізує утворення ковалентного зв'язку фосфодіефіру між 3'-OH кінцем одного фрагмента ДНК та 5-футовим фосфатним кінцем іншого фрагмента, стабілізації цукрово-фосфатного хребта молекули ДНК.
Припинення
Після того, як повна хромосома буде реплікована, має відбутися припинення реплікації ДНК. Хоча про ініціювання реплікації відомо багато, про процес припинення відомо менше. Після реплікації отримані повні кругові геноми прокаріотів об'єднуються, що означає, що кругові хромосоми ДНК зблоковані і повинні бути відокремлені один від одного. Це досягається завдяки активності бактеріальної топоізомерази IV, яка вводить дволанцюгові розриви в молекули ДНК, дозволяючи їм відокремлюватися один від одного; фермент потім повторює кругові хромосоми. Дозвіл конкатемерів - це питання, унікальне для реплікації прокаріотичної ДНК через їх кругові хромосом. Оскільки як бактеріальна ДНК-гіраза, так і топоізомераза IV відрізняються від своїх еукаріотичних аналогів, ці ферменти служать мішенями для класу антимікробних препаратів, які називаються хінолонами.
| Фермент або фактор | Функція |
|---|---|
| ДНК-пул I | Екзонуклеазна активність видаляє праймер РНК і замінює його новосинтезованою ДНК |
| ДНК-пол III | Основний фермент, який додає нуклеотиди в напрямку 5' до 3' |
| Хелікейс | Відкриває спіраль ДНК шляхом розриву водневих зв'язків між азотистими основами |
| Лігаза | Заклеює проміжки між фрагментами Окадзакі на відстаючої пасма, щоб створити одну суцільну нитку ДНК |
| Примаза | Синтезує праймери РНК, необхідні для початку реплікації |
| Одножильні зв'язуючі білки | Зв'язуйтеся з одноцепочечной ДНК, щоб запобігти водневому зв'язку між нитками ДНК, реформуючи дволанцюгову ДНК |
| Розсувний затискач | Допомагає утримувати ДНК pol III на місці, коли нуклеотиди додаються |
| Топоізомераза II (ДНК-гіраза) | Розслаблює суперспіральну хромосому, щоб зробити ДНК більш доступною для ініціації реплікації; допомагає зняти стрес на ДНК при розмотуванні, викликаючи розриви, а потім перегерметизуючи ДНК |
| Топоізомераза IV | Вводить однонитковий розрив на конкатеновані хромосоми, щоб звільнити їх один від одного, а потім повторює ДНК |
Вправа\(\PageIndex{2}\)
- Який фермент розриває водневі зв'язки, що утримують дві нитки ДНК разом, так що може відбутися реплікація?
- Це відстаюча пасмо або провідна пасмо, яка синтезується в напрямку до відкриття вилки реплікації?
- Який фермент відповідає за видалення праймерів РНК у нещодавно реплікованої бактеріальної ДНК?
Реплікація ДНК у еукаріотів
Еукаріотичні геноми набагато складніші і більші, ніж прокаріотичні геноми, і зазвичай складаються з множинних лінійних хромосом (табл.\(\PageIndex{2}\)). Наприклад, геном людини має 3 мільярди пар основ на гаплоїдний набір хромосом, а 6 мільярдів пар основ вставляються під час реплікації. Існує кілька джерел реплікації на кожній еукаріотичної хромосомі (рис.\(\PageIndex{5}\)); геном людини має від 30 000 до 50 000 походження реплікації. Швидкість реплікації становить приблизно 100 нуклеотидів в секунду - 10 разів повільніше, ніж реплікація прокаріотів.
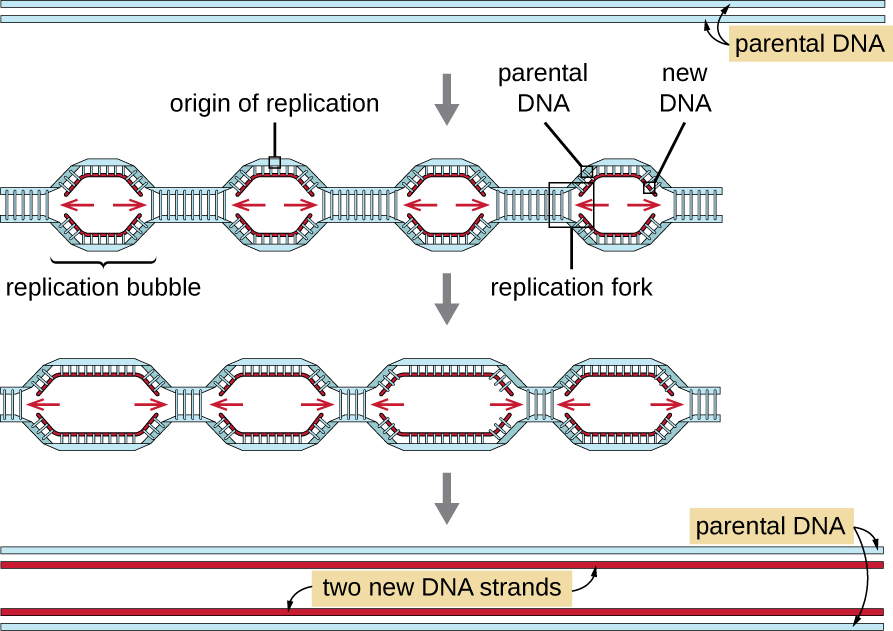
Істотні етапи реплікації у еукаріотів такі ж, як і у прокаріотів. Перш ніж реплікація може початися, ДНК повинна бути доступна як шаблон. Еукаріотична ДНК сильно перекручена і упакована, чому сприяють багато білків, включаючи гістони (див. Структура і функція клітинних геномів). За походженням реплікації комплекс пререплікації, що складається з декількох білків, включаючи геліказу, утворює та набирає інші ферменти, що беруть участь у ініціюванні реплікації, включаючи топоізомеразу для розслаблення суперколінгу, одноцепочечного зв'язуючого білка, РНК-примази та ДНК-полімерази. Після початку реплікації, в процесі, подібному до того, що виявлено у прокаріотів, подовженню сприяють еукаріотичні ДНК-полімерази. Провідна нитка безперервно синтезується ензимом еукаріотичної полімерази pol δ, в той час як відстаюча пасмо синтезується pol ε. Білок ковзного затиску утримує ДНК-полімеразу на місці, щоб вона не відпадала з ДНК. Фермент рибонуклеаза Н (РНАза Н) замість ДНК-полімерази, як у бактерій, видаляє праймер РНК, який потім замінюється нуклеотидами ДНК. Прогалини, які залишилися, герметизуються ДНК-лігазою.
Оскільки еукаріотичні хромосоми є лінійними, можна очікувати, що їх реплікація буде більш простою. Як і у прокаріотів, еукаріотична ДНК-полімераза може додавати нуклеотиди лише в напрямку 5' до 3'. У провідній пасмі синтез триває до тих пір, поки не досягне або кінця хромосоми, або іншої вилки реплікації, що прогресує в протилежному напрямку. На відстає пасма синтезується ДНК короткими відрізками, кожен з яких ініціюється окремим праймером. Коли вилка реплікації досягає кінця лінійної хромосоми, ніде зробити праймер для фрагмента ДНК, який буде скопійований в кінці хромосоми. Таким чином, ці кінці залишаються непарними, і з часом вони можуть ставати поступово коротшими, оскільки клітини продовжують ділитися.
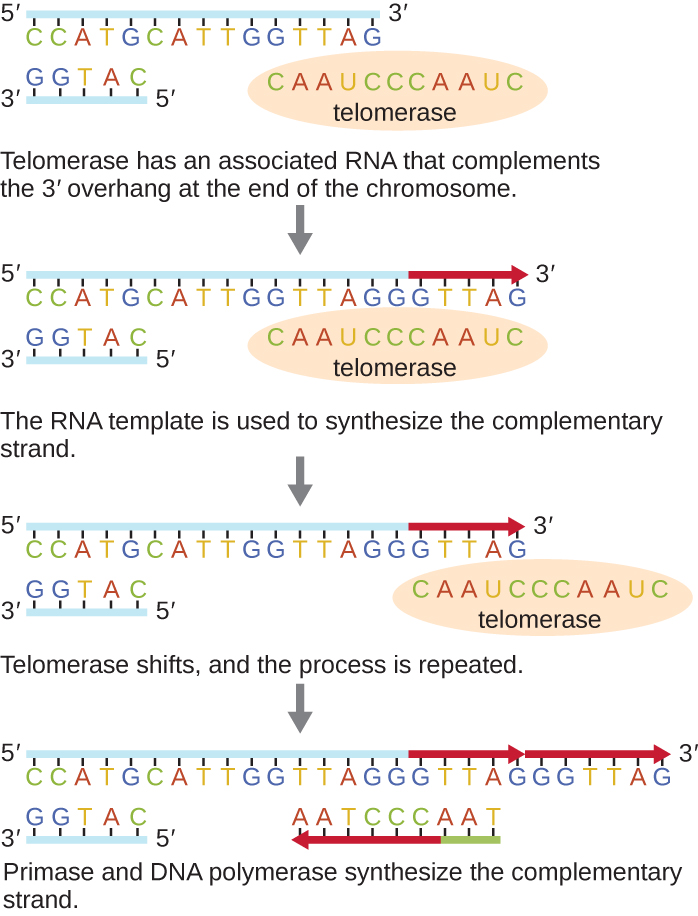
Кінці лінійних хромосом відомі як теломери і складаються з некодуючих повторюваних послідовностей. Теломери захищають послідовності кодування від втрати, оскільки клітини продовжують ділитися. У людей послідовність шести базових пар, TTAGGG, повторюється від 100 до 1000 разів, щоб сформувати теломер. Відкриття ферменту теломерази (рис.\(\PageIndex{6}\)) прояснило наше розуміння того, як підтримуються кінці хромосоми. Теломераза містить каталітичну частину і вбудований шаблон РНК. Він прикріплюється до кінця хромосоми, а додаткові основи до шаблону РНК додаються на 3' кінці ланцюга ДНК. Після того, як 3-футовий кінець шаблону відстаючого пасма досить витягнутий, ДНК-полімераза може додати нуклеотиди, що доповнюють кінці хромосом. Таким чином відбувається реплікація кінців хромосом. У людей теломераза, як правило, активна в статевих клітині і стовбурових клітині дорослих; вона не активна в соматичних клітині дорослих і може бути пов'язана зі старінням цих клітин. Еукаріотичні мікроби, включаючи гриби та найпростіші, також виробляють теломеразу для підтримки цілісності хромосом. За відкриття теломерази та її дії Елізабет Блекберн (1948—) отримала Нобелівську премію з медицини та фізіології в 2009 році.
| Нерухомість | Бактерії | Еукаріот |
|---|---|---|
| будова геному | Одинарна кругова хромосома | Множинні лінійні хромосоми |
| Кількість походження на хромосому | Одномісний | Множинні |
| Швидкість реплікації | 1000 нуклеотидів в секунду | 100 нуклеотидів в секунду |
| Теломераза | Ні | Подарунок |
| Видалення грунтовки РНК | ДНК-пул I | Рнза H |
| подовження пасом | ДНК-пол III | пол δ, пол ε |
Вправа\(\PageIndex{3}\)
- Чим відрізняється походження реплікації між еукаріотами і прокаріотами?
- Які ферменти полімерази відповідають за синтез ДНК при еукаріотичної реплікації?
- Що виявляється на кінцях хромосом у еукаріот і чому?
Реплікація ДНК екстрахромосомних елементів: плазміди та віруси
Для копіювання своїх нуклеїнових кислот, плазміди та віруси часто використовують варіації шаблону реплікації ДНК, описані для геномів прокаріотів. Для отримання додаткової інформації про широкий спектр стратегій реплікації вірусів див. Життєвий цикл вірусу.
Коло кочення реплікації
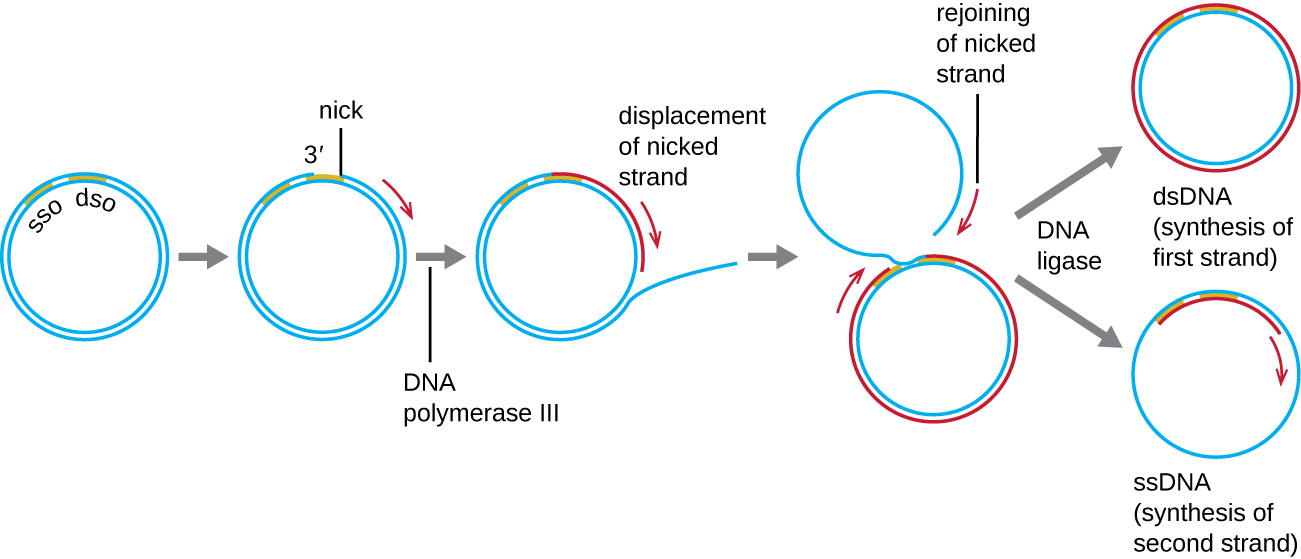
Тоді як багато бактеріальних плазмід (див. Унікальні характеристики прокаріотичних клітин) відтворюються процесом, подібним до того, який використовується для копіювання бактеріальної хромосоми, інших плазмід, декількох бактеріофагів та деяких вірусів еукаріотів використовують реплікацію кочення кола (рис.\(\PageIndex{7}\)). Циркулярний характер плазмід і циркуляризація деяких вірусних геномів при зараженні роблять це можливим. Реплікація прокатного кола починається з ферментативного защемлення однієї нитки двожильної кругової молекули на місці двожильного походження (dso). У бактерій ДНК-полімераза III зв'язується з групою 3'-OH вирізаної нитки і починає однонаправлено тиражувати ДНК, використовуючи невирізану нитку як шаблон, витісняючи нитку, як це робить. Завершення реплікації ДНК на місці вихідного ніка призводить до повного зміщення вирізаної нитки, яка потім може рециркуляризуватися в одноцепочечную молекулу ДНК. РНК примаза потім синтезує праймер для ініціювання реплікації ДНК на одноланцюговому місці походження (sso) молекули одноцепочечной ДНК (ssDNA), в результаті чого дволанцюгова молекула ДНК (dsDNA) ідентична іншій круговій молекулі ДНК.
Вправа\(\PageIndex{4}\)
Чи є відстаюча пасмо в коченому колі реплікації? Чому чи чому ні?
Ключові поняття та резюме
- Процес реплікації ДНК є напівконсервативним, в результаті чого дві молекули ДНК, кожна з яких має одну батьківську нитку ДНК і одну знову синтезовану нитку.
- У бактерій ініціювання реплікації відбувається у походження реплікації, де суперспіральна ДНК розмотується ДНК-гіразою, зроблена однониткової геліказою, і пов'язана однонитковим зв'язуючим білком для підтримки його одно- багатожильний стан. Примаза синтезує короткий праймер РНК, забезпечуючи вільну 3'-OH групу, до якої ДНК-полімераза III може додавати нуклеотиди ДНК.
- Під час подовження провідна нитка ДНК синтезується безперервно з одного праймера. Відстаюча пасмо синтезується розривно короткими фрагментами Окадзакі, кожен вимагає свого праймера. РНК-праймери видаляються і замінюються ДНК-нуклеотидами бактеріальною ДНК-полімеразою I, а ДНК-лігаза ущільнює проміжки між цими фрагментами.
- Припинення реплікації у бактерій передбачає дозвіл кругових конкатемерів ДНК топоізомеразою IV для звільнення двох копій кругової хромосоми.
- Еукаріоти зазвичай мають кілька лінійних хромосом, кожна з яких має множинне походження реплікації. В цілому реплікація у еукаріотів подібна до реплікації у прокаріотів.
- Лінійний характер еукаріотичних хромосом обумовлює необхідність теломерів для захисту генів поблизу кінця хромосом. Теломераза розширює теломери, запобігаючи їх деградації, в деяких типах клітин.
- Реплікація коченого кола - це тип швидкого однонаправленого синтезу ДНК кругової молекули ДНК, що використовується для реплікації деяких плазмід.