10.2: Будова та функція ДНК
- Page ID
- 3896

Цілі навчання
- Опишіть біохімічну будову дезоксирибонуклеотидів
- Визначте пари основ, що використовуються при синтезі дезоксирибонуклеотидів
- Поясніть, чому подвійна спіраль ДНК описується як антипаралельна
У Мікробному метаболізмі ми обговорили три класи макромолекул: білки, ліпіди та вуглеводи. У цьому розділі ми обговоримо четвертий клас макромолекул: нуклеїнові кислоти. Як і інші макромолекули, нуклеїнові кислоти складаються з мономерів, званих нуклеотидами, які полімеризуються з утворенням великих ниток. Кожна нитка нуклеїнової кислоти містить певні нуклеотиди, які з'являються в певному порядку всередині пасма, звані її базовою послідовністю. Базова послідовність дезоксирибонуклеїнової кислоти (ДНК) відповідає за перенесення та збереження спадкової інформації в клітині. У «Механізмах мікробної генетики» ми детально обговоримо способи, якими ДНК використовує власну базову послідовність для направлення власного синтезу, а також синтез РНК і білків, що, в свою чергу, породжує продукти з різноманітною структурою і функцією. У цьому розділі ми обговоримо основну будову і функції ДНК.
Нуклеотиди ДНК
Будівельні блоки нуклеїнових кислот - нуклеотиди. Нуклеотиди, що складають ДНК, називаються дезоксирибонуклеотидами. Три компоненти дезоксирибонуклеотиду - це п'ятивуглецевий цукор, який називається дезоксирибозою, фосфатною групою та азотиста основа, азотовмісна кільцева структура, яка відповідає за додаткове сполучення основи між нитками нуклеїнової кислоти (рис.\(\PageIndex{1}\)). Атоми вуглецю п'ятивуглецевої дезоксирибози пронумеровані 1', 2', 3', 4' і 5' (1' читається як «один простий»). Нуклеозид містить п'ятивуглецевий цукор та азотисту основу.
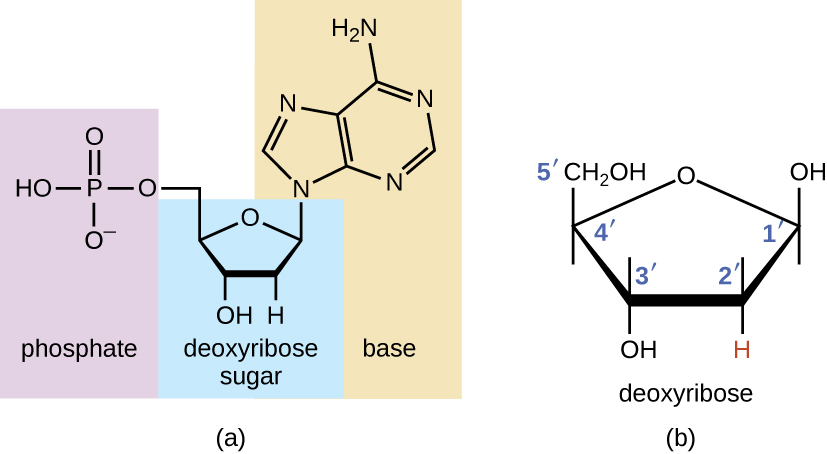
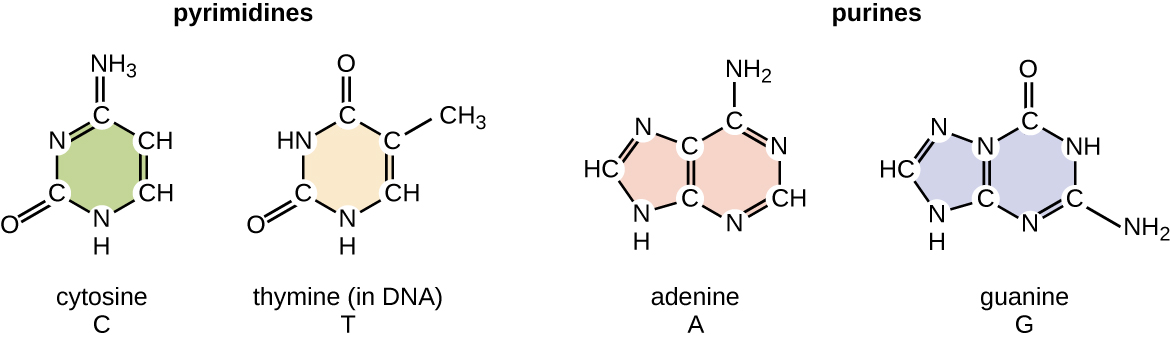
Дезоксирибонуклеотид названий відповідно до азотистих підстав (рис.\(\PageIndex{2}\)). Азотисті основи аденін (А) і гуанін (G) є пуринами; вони мають подвійну кільцеву структуру з шестивуглецевим кільцем, сплавленим до п'ятивуглецевого кільця. Піримідини, цитозин (С) і тимін (Т) - це менші азотисті підстави, які мають лише шестивуглецеву кільцеву структуру.
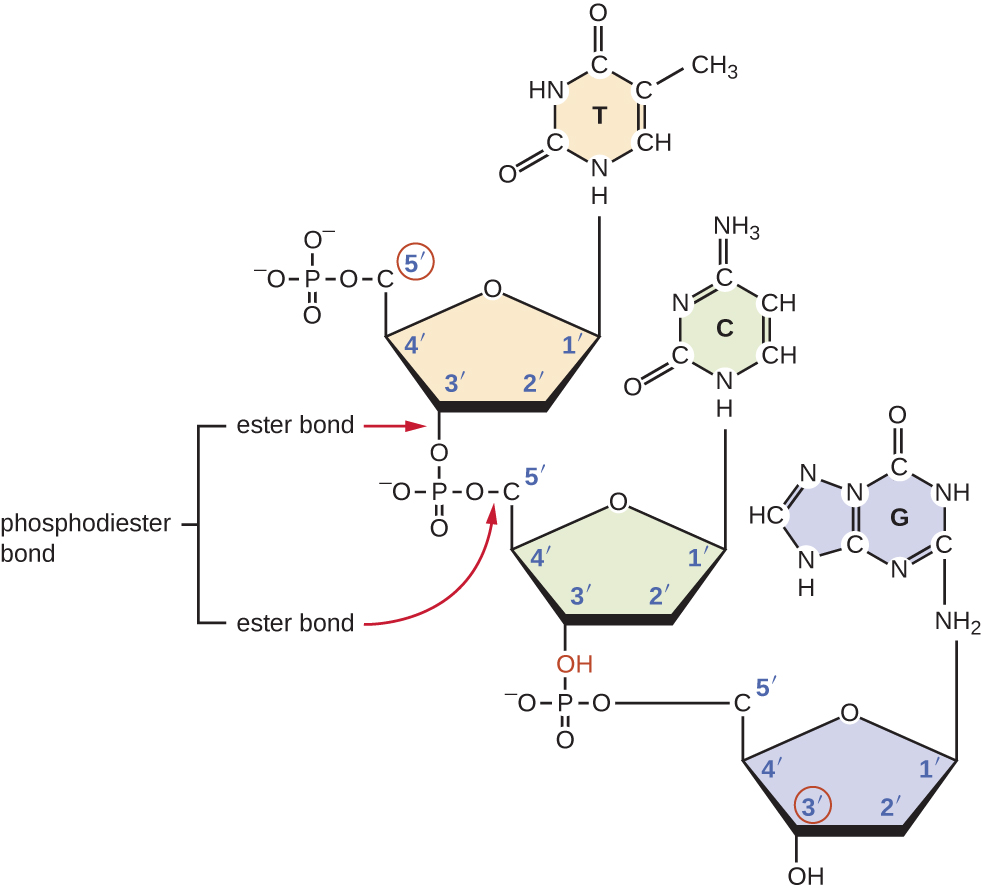
Окремі нуклеозидні трифосфати поєднуються між собою ковалентними зв'язками, відомими як 5'-3' фосфодіефірні зв'язки, або зв'язками, за допомогою яких фосфатна група приєднується до 5' вуглецю цукру одного нуклеотидного зв'язку з гідроксильною групою 3' вуглецю цукру наступного нуклеотиду. Фосфодіефірний зв'язок між нуклеотидами утворює цукрово-фосфатну кістку, що чергується цукрово-фосфатна структура, що становить каркас нитка нуклеїнової кислоти (рис.\(\PageIndex{3}\)). В процесі полімеризації використовуються дезоксинуклеотидні трифосфати (dnTp). Для побудови цукрово-фосфатної магістралі два кінцеві фосфати виділяються з dNTP у вигляді пірофосфату. Отримана нитка нуклеїнової кислоти має вільну фосфатну групу на 5'вуглецевому кінці і вільну гідроксильну групу на 3' вуглецевому кінці. Дві невикористані фосфатні групи з нуклеотидного трифосфату виділяються у вигляді пірофосфату під час формування фосфодіефірних зв'язків. Пірофосфат згодом гідролізується, вивільняючи енергію, яка використовується для приводу полімеризації нуклеотидів.
Вправа\(\PageIndex{1}\)
Що мається на увазі під кінцями 5' і 3' нуклеїнової кислоти?
Відкриття подвійної спіралі
До початку 1950-х років накопичилися значні докази, що свідчать про те, що ДНК є генетичним матеріалом клітин, і тепер гонка продовжувала виявляти її тривимірну структуру. Приблизно в цей час австрійський біохімік Ервін Чаргафф 1 (1905—2002) дослідив вміст ДНК у різних видів і виявив, що аденін, тимін, гуанін та цитозин не були виявлені в рівних кількостях, і що він варіювався від виду до виду, але не між особинами однакових види. Він виявив, що кількість аденіну дуже близька до зрівнювання кількості тиміну, а кількість цитозину була дуже близькою до зрівнювання кількості гуаніну, або A = T і G = C. Ці відносини також відомі як правила Чаргаффа.
Інші вчені також активно досліджували цю галузь протягом середини 20 століття. У 1952 році американський вчений Лінус Полінг (1901—1994) був провідним світовим хіміком-структурним і непарним фаворитом для вирішення структури ДНК. Полінг раніше відкрив структуру білкових α спіралей, використовуючи рентгенівську дифракцію, і на основі рентгенівських дифракційних зображень ДНК, зроблених у його лабораторії, він запропонував триланцюжкову модель ДНК. 2 У той же час британські дослідники Розалінда Франклін (1920—1958) та її аспірант Р.Г. Гослінг також використовували рентгенівську дифракцію для розуміння структури ДНК (рис.\(\PageIndex{4}\)). Саме наукова експертиза Франкліна призвела до отримання більш чітко визначених рентгенівських дифракційних зображень ДНК, які чітко показували б загальну структуру ДНК з подвійною спіраллю.

Джеймс Уотсон (1928—), американський вчений, та Френсіс Крик (1916—2004), британський вчений, працювали разом у 1950-х роках над виявленням структури ДНК. Вони використовували правила Чаргаффа та рентгенівські дифракційні зображення Франкліна та Вілкінса волокон ДНК, щоб з'єднати пурин-піримідинове сполучення подвійної спіральної молекули ДНК (рис.\(\PageIndex{5}\)). У квітні 1953 року Уотсон і Крик опублікували свою модель подвійної спіралі ДНК в Nature. 3 Це ж питання додатково включало статті Уілкінса та колег, 4, а також Франкліна та Гослінга, 5 кожна з яких описує різні аспекти молекулярної структури ДНК. У 1962 році Джеймс Уотсон, Френсіс Крик та Моріс Вілкінс були удостоєні Нобелівської премії з фізіології та медицини. На жаль, на той час Франклін помер, а Нобелівські премії в той час не були присуджені посмертно. Продовжувалася робота, однак, над вивченням структури ДНК. У 1973 році Олександр Річ (1924—2015) та його колеги змогли проаналізувати кристали ДНК для підтвердження та подальшого з'ясування структури ДНК. 6

Вправа\(\PageIndex{2}\)
Які вчені отримують більшу частину заслуги за опис молекулярної структури ДНК?
Структура ДНК
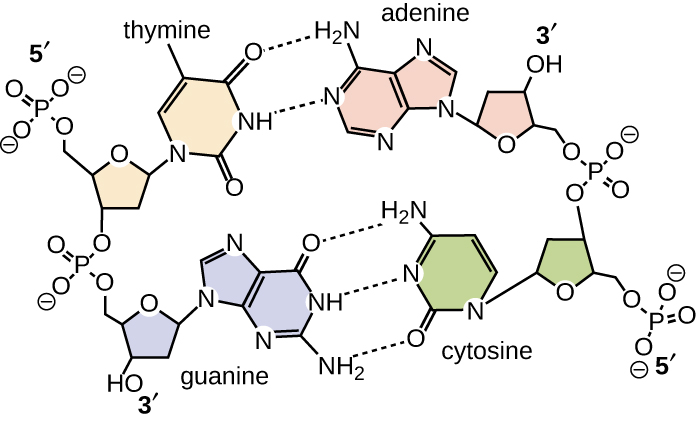
Уотсон і Крик запропонували, щоб ДНК складалася з двох ниток, які скручені навколо один одного, утворюючи правосторонню спіраль. Дві нитки ДНК є антипаралельними, таким чином, що 3' кінець однієї нитки звернений до 5' кінця іншої (рис.\(\PageIndex{6}\)). 3' кінець кожного пасма має вільну гідроксильну групу, тоді як 5' кінець кожної нитки має вільну фосфатну групу. Цукор і фосфат полімеризованих нуклеотидів утворюють кістяк структури, тоді як азотисті підстави укладаються всередину. Ці азотисті підстави на внутрішній стороні молекули взаємодіють один з одним, спарюючи основу.
Аналіз дифракційних закономірностей ДНК визначив, що в ДНК існує приблизно 10 основ. Асиметричний інтервал цукрово-фосфатних магістралей утворює великі канавки (де хребет знаходиться далеко один від одного) і незначні канавки (де хребет знаходиться близько один до одного) (рис.\(\PageIndex{6}\)). Ці канавки - це місця, де білки можуть зв'язуватися з ДНК. Зв'язування цих білків може змінювати структуру ДНК, регулювати реплікацію або регулювати транскрипцію ДНК в РНК.
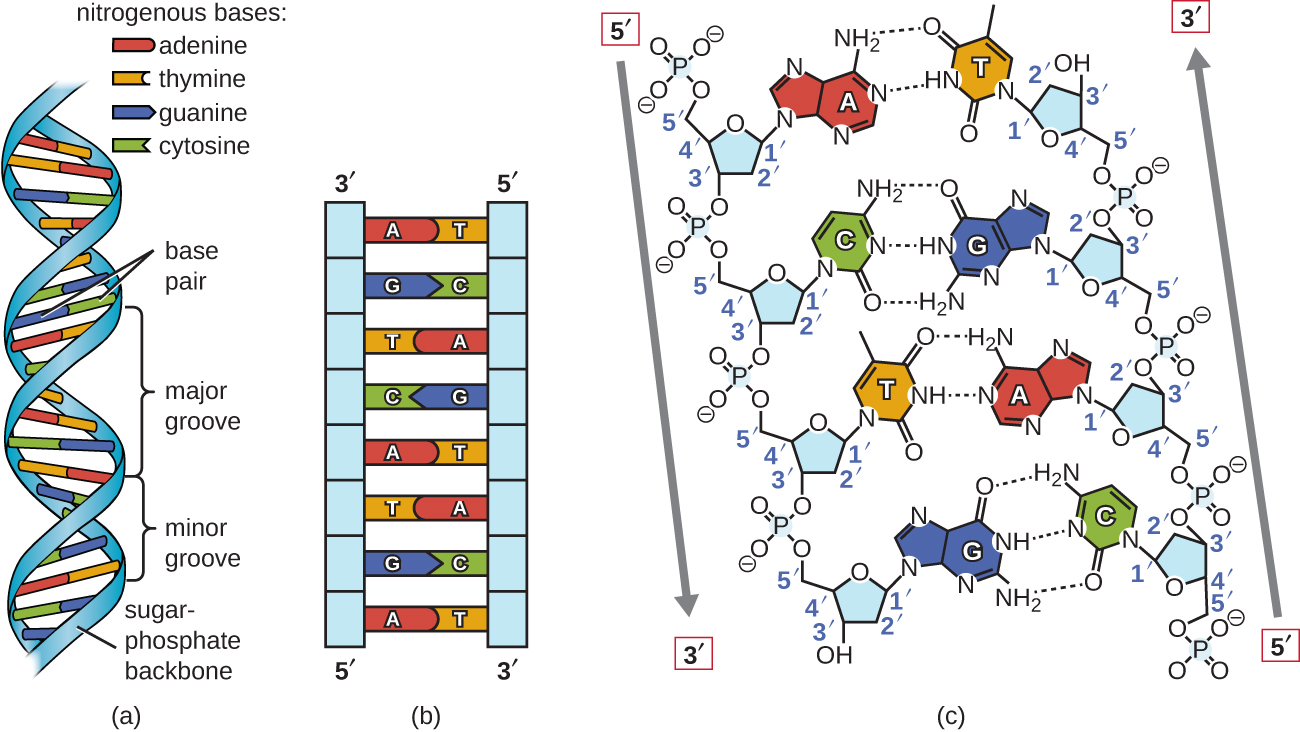
Спарювання основи відбувається між пурином і піримідином. У ДНК аденін (А) і тимін (Т) є комплементарними парами основ, а цитозин (С) і гуанін (G) також є комплементарними парами основ, пояснюючи правила Чаргаффа (рис.\(\PageIndex{7}\)). Пари основи стабілізуються водневими зв'язками; аденін і тимін утворюють між ними дві водневі зв'язки, тоді як цитозин і гуанін утворюють між ними три водневі зв'язки.
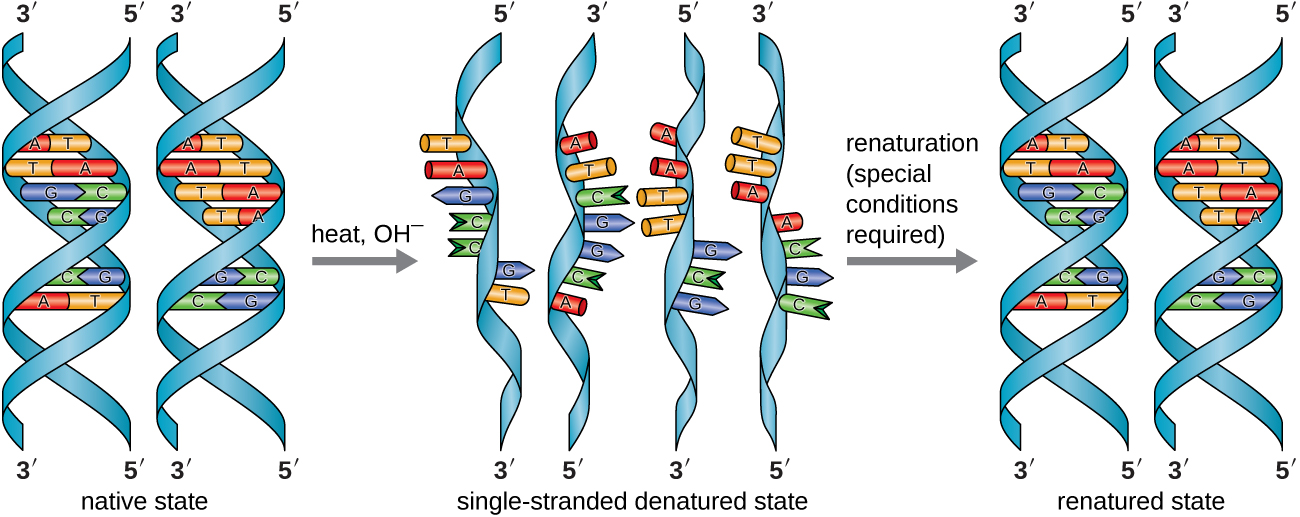
У лабораторії, піддаючи дві нитки ДНК подвійної спіралі високим температурам або певним хімічним речовинам, може розірвати водневі зв'язки між комплементарними основами, розділяючи таким чином нитки на дві окремі окремі нитки ДНК (одноцепочечной ДНК [SsDNA]). Цей процес називається денатурацією ДНК і є аналогом денатурації білка, як описано в розділі Білки. Нитки SSDNA також можуть бути складені разом як дволанцюгова ДНК (dsDNA), шляхом відпалу або ренатурування шляхом охолодження або видалення хімічних денатурантів, що дозволяє цим водневим зв'язкам реформуватися. Здатність штучно маніпулювати ДНК таким способом є основою декількох важливих методик в біотехнології (рис.\(\PageIndex{8}\)). Через додатковий водневий зв'язок між базовою парою C = G ДНК з високим вмістом ГК важче денатурувати, ніж ДНК з меншим вмістом ГК.
Перегляньте анімацію про структуру ДНК з навчального центру ДНК, щоб дізнатися більше.
Вправа\(\PageIndex{3}\)
Що таке дві комплементарні базові пари ДНК і як вони пов'язані між собою?
Функція ДНК
ДНК зберігає інформацію, необхідну для побудови та контролю клітини. Передача цієї інформації від матері до дочірніх клітин називається вертикальним переносом генів і відбувається через процес реплікації ДНК. ДНК реплікується, коли клітина робить дублікат своєї ДНК, потім клітина ділиться, в результаті чого відбувається правильний розподіл однієї копії ДНК на кожну результуючу клітину. ДНК також може бути ферментативно деградована і використовуватися як джерело нуклеозидів і нуклеотидів для клітини. На відміну від інших макромолекул, ДНК не виконує структурної ролі в клітині.
Вправа\(\PageIndex{4}\)
Як ДНК передає генетичну інформацію потомству?
Прокладаючи шлях для жінок у науці та медичних професіях
Історично жінки були недостатньо представлені в науках і медицині, і часто їх новаторські внески залишаються відносно непоміченими. Наприклад, хоча Розалінда Франклін проводила рентгенівські дифракційні дослідження, що демонструють подвійну гвинтову структуру ДНК, саме Уотсон і Крик прославилися цим відкриттям, спираючись на її дані. Все ще залишаються великі суперечки щодо того, чи було їх придбання її даних доречним та чи сприяли конфлікти особистості та гендерні упередження затриманому визнанню її значних внесків. Аналогічно, Барбара МакКлінток робила новаторську роботу в генетиці кукурудзи (кукурудзи) з 1930-х по 1950-х, виявивши транспозони (стрибаючі гени), але вона не була визнана набагато пізніше, отримавши Нобелівську премію з фізіології або медицини в 1983 році (рис.\(\PageIndex{9}\)).
Сьогодні жінки все ще залишаються недопредставленими в багатьох галузях науки і медицини. Хоча більше половини ступенів бакалаврату в галузі науки присуджуються жінкам, лише 46% докторських ступенів в науці присуджуються жінкам. В академічних колах кількість жінок на кожному рівні кар'єрного зростання продовжує зменшуватися, причому жінки займають менше третини посад вчених рівня доктора філософії на посаді, а також менше чверті повних професорських посад в 4-річних коледжах та університетах. 7 Навіть у медичних професіях, як і майже у всіх інших сферах, жінки часто недопредставлені у багатьох медичних кар'єрах і заробляють значно менше, ніж їхні колеги-чоловіки, як показано в дослідженні 2013 року, опублікованому журналом Американської медичної асоціації. 8
Чому такі розбіжності продовжують існувати і як ми порушуємо ці цикли? Ситуація складна і, ймовірно, є результатом поєднання різних факторів, включаючи те, як суспільство обумовлює поведінку дівчат з юного віку та підтримує їх інтереси, як професійно, так і особисто. Деякі припустили, що жінки не належать до лабораторії, включаючи лауреата Нобелівської премії Тіма Ханта, чиї громадські коментарі 2015 року свідчать про те, що жінки занадто емоційні для науки 9 були зустрінуті з широким засудженням.
Можливо, дівчат варто більше підтримувати з юних років в областях науки і математики (рис.\(\PageIndex{9}\)). Програми науки, техніки, техніки та математики (STEM), спонсоровані Американською асоціацією жінок-університетів (AAUW) 10 та Національним управлінням з аеронавтики та космічного простору (NASA) 11, є чудовими прикладами програм, які пропонують таку підтримку. Внески жінок у науку повинні бути більш широко відомі громадськості, а маркетинг, орієнтований на молодих дівчат, повинен включати більше образів історично та професійно успішних жінок-вчених та медичних працівників, заохочуючи всіх світлих молодих умів, включаючи дівчат та жінок, до переслідування кар'єра в науці та медицині.

Клінічна спрямованість: Частина 2
Грунтуючись на його симптомах, лікар Алекса підозрює, що він страждає від харчової хвороби, яку він придбав під час подорожей. Можливості включають бактеріальну інфекцію (наприклад, ентеротоксигенну кишкову паличку, холерний вібріон, Campylobacter jejuni, Salmonella), вірусну інфекцію (ротавірус або норовірус) або найпростішу інфекцію (лямблії, Cryptosporidium parvum , Або Ентамоеба гістолітика).
Його лікар замовляє зразок калу для виявлення можливих збудників (наприклад, бактерій, кіст) та пошуку наявності крові, оскільки певні типи інфекційних агентів (наприклад, C. jejuni, Salmonella та E. histolytica) пов'язані з виробленням кров'яних табурети.
Зразок калу Алекса не показав ні крові, ні кісти. Після аналізу зразка калу та на основі його недавньої історії подорожей лікар лікарні запідозрив, що Алекс страждає від діареї мандрівника, спричиненої ентеротоксигенною кишковою паличкою (ETEC), збудником діареї більшості мандрівників. Щоб перевірити діагноз та виключити інші можливості, лікар Алекса замовив діагностичний лабораторний тест його зразка калу для пошуку послідовностей ДНК, що кодують специфічні фактори вірулентності ETEC. Лікар доручив Алексу пити багато рідини, щоб замінити те, що він втратив, і виписав його з лікарні.
ETEC виробляє кілька факторів вірулентності, кодованих плазмою, які роблять його патогенним порівняно з типовою кишковою паличкою. До них відносяться виділяються токсини теплолабільний ентеротоксин (ЛТ) і теплостійкий ентеротоксин (ST), а також фактор колонізації (CF). І LT, і ST викликають виведення іонів хлориду з клітин кишечника в просвіт кишечника, викликаючи подальшу втрату води з клітин кишечника, що призводить до діареї. CF кодує бактеріальний білок, який допомагає бактерії прилипати до слизової оболонки тонкої кишки.
Вправа\(\PageIndex{5}\)
Чому лікар Алекса використовував генетичний аналіз замість виділення бактерій із зразка калу або прямого фарбування Грама лише зразка калу?
Ключові поняття та резюме
- Нуклеїнові кислоти складаються з нуклеотидів, кожен з яких містить пентозний цукор, фосфатну групу і азотисту основу. Дезоксирибонуклеотиди в ДНК містять дезоксирибозу як пентозу цукру.
- ДНК містить піримідини цитозин і тимін, а також пурини аденін і гуанін.
- Нуклеотиди пов'язані між собою фосфодіефірними зв'язками між 5'фосфатною групою одного нуклеотиду і 3' гідроксильною групою іншого. Нитка нуклеїнової кислоти має вільну фосфатну групу на кінці 5' і вільну гідроксильну групу на 3' кінці.
- Чаргафф виявив, що кількість аденіну приблизно дорівнює кількості тиміну в ДНК, і що кількість гуаніну приблизно дорівнює цитозину. Пізніше ці відносини були визначені як обумовлені взаємодоповнюючим базовим сполученням.
- Уотсон і Крик, спираючись на роботу Чаргаффа, Франкліна і Гослінга та Вілкінса, запропонували модель подвійної спіралі та сполучення основи для структури ДНК.
- ДНК складається з двох взаємодоповнюючих ниток, орієнтованих антипаралельно один одному з магістралями фосфодіефіру на зовнішній стороні молекули. Азотисті підстави кожної пасма звернені один до одного і доповнюють собою підстави водню зв'язуються між собою, стабілізуючи подвійну спіраль.
- Тепло або хімічні речовини можуть порушити водневі зв'язки між комплементарними основами, денатуруючи ДНК. Охолодження або видалення хімічних речовин може призвести до ренатурації або повторного відпалу ДНК, дозволяючи водневим зв'язкам реформуватися між додатковими основами.
- ДНК зберігає інструкції, необхідні для побудови та контролю клітини. Ця інформація передається від батьків до потомства через вертикальний перенос генів.
Виноски
- 1 Н. Кресге та ін. «Правила Чаргаффа: Робота Ервіна Чаргаффа». Журнал біологічної хімії 280 (2005) :e21.
- 2 Л. Паулінг, «Пропонована структура нуклеїнових кислот.» Праці Національної академії наук Сполучених Штатів Америки 39 № 2 (1953) :84—97.
- 3 Джей Д.Д. Ватсон, Ф.К. «Структура дезоксирибози нуклеїнової кислоти». Природа 171 № 4356 (1953) :737—738.
- 4 М.Х.Ф. Уілкінс та ін. «Молекулярна структура дезоксипентозних нуклеїнових кислот». Природа 171 № 4356 (1953) :738—740.
- 5 Р.Франклін, Р.Г. Гослінг. «Молекулярна конфігурація в тимоноклеат натрію». Природа 171 № 4356 (1953) :740—741.
- 6 RO День та ін. «Кристалічний фрагмент подвійної спіралі: структура динуклеозидного фосфату гуаніліл-3',5'-цітидину». Праці Національної академії наук Сполучених Штатів Америки 70 № 3 (1973) :849—853.
- 7 Н.Г. Вольфінгер «Для жінок-вчених немає гарного часу, щоб мати дітей». Атлантичний 29 липня 2013. www.theatlantic.com/sexes/arc... ілдрен/278165/.
- 8 С.А. Сібері та ін. «Тенденції заробітку чоловіків і жінок медичних працівників у Сполучених Штатах, 1987 по 2010 рік». Журнал Американської медичної асоціації внутрішньої медицини 173 № 18 (2013) :1748—1750.
- 9 Е. Чунг. «Тім Хант, сексизм і наука: справжні «неприємності з дівчатами» в лабораторіях». CBC Новини Технології та Наука, 12 червня 2015 року. http://www.cbc.ca/news/technology/ti...labs-1.3110133. Доступ до 04.08.2016.
- 10 Американська асоціація жінок-університетів. «Побудова трубопроводу STEM для дівчат та жінок». www.aauw.org/what-we-do/stem-education/. Доступ до 10 червня 2016 року.
- 11 Національне управління з аеронавтики та космічного простору. «Просвітницькі програми: Ініціатива жінок та дівчат». http://women.nasa.gov/outreach-programs/. Доступ до 10 червня 2016 року.