10.1: Використання мікробіології для розкриття таємниць життя
- Page ID
- 3899

Цілі навчання
- Опишіть відкриття нуклеїнової кислоти і нуклеотидів
- Поясніть історичні експерименти, які привели до характеристики ДНК
- Опишіть, як мікробіологія та мікроорганізми були використані для виявлення біохімії генів
- Поясніть, як вчені встановили зв'язок між ДНК і спадковістю
Клінічна спрямованість: Частина 1
Алекс - 22-річний студент коледжу, який відпочивав у Пуерто-Вальярта, Мексика, на весняні канікули. На жаль, через два дні після польоту додому в Огайо він почав відчувати спазми в животі і велику водянисту діарею. Через свій дискомфорт він звернувся за медичною допомогою до великої лікарні Цинциннаті неподалік.
Вправа\(\PageIndex{1}\)
Які види інфекцій або інші стани можуть бути відповідальними?
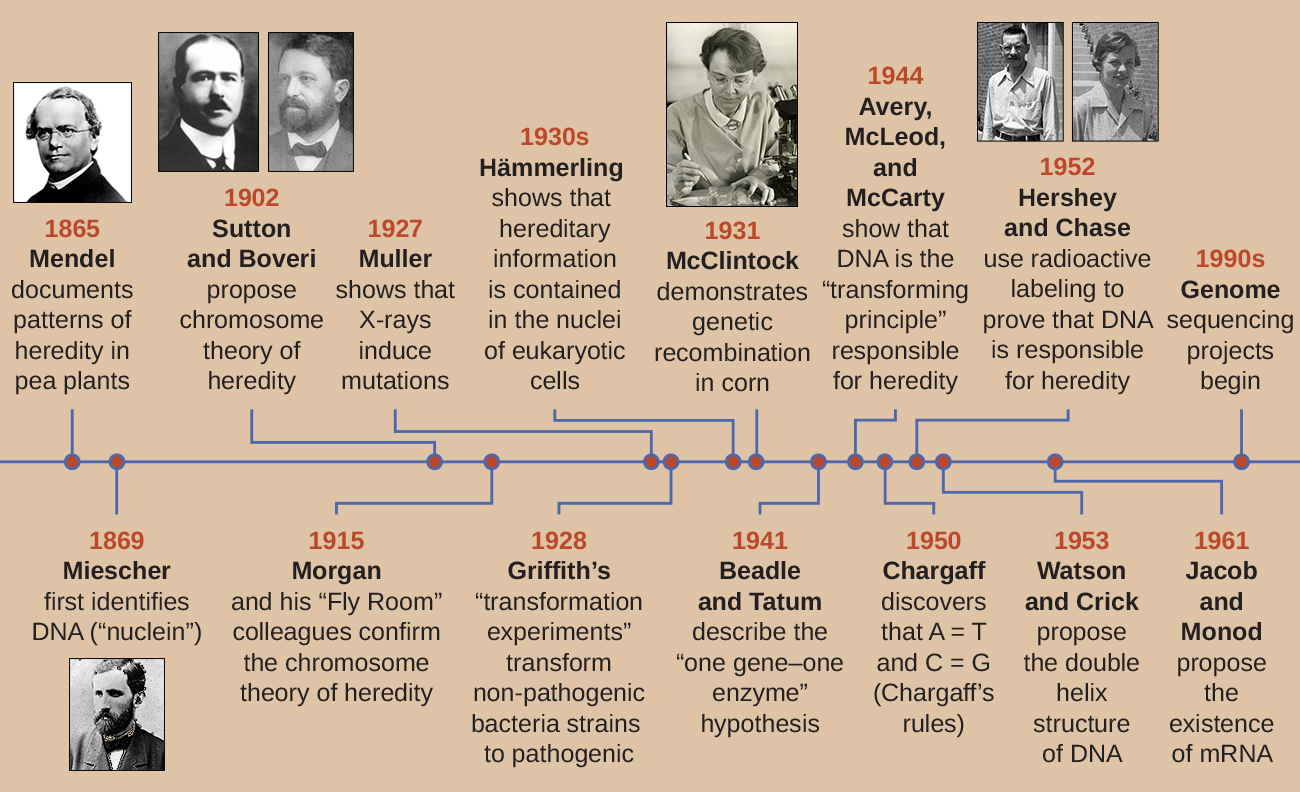
На початку 20 століття ДНК ще не визнавалася генетичним матеріалом, що відповідає за спадковість, перехід ознак з одного покоління в інше. Насправді значна частина досліджень була відкинута до середини 20 століття. Наукове співтовариство неправильно вважало, що процес успадкування передбачає змішування батьківських рис, що породило проміжний фізичний вигляд у потомства; цей гіпотетичний процес виявився правильним через те, що ми знаємо зараз як безперервну зміну, що є результатом дії багатьох генів, щоб визначити ту чи іншу характеристику, як людський зріст. Потомство, здається, є «сумішшю» рис їхніх батьків, коли ми дивимось на характеристики, які демонструють безперервні зміни. Теорія змішування успадкування стверджувала, що початкові батьківські риси були втрачені або поглинені змішуванням у потомстві, але тепер ми знаємо, що це не так.
Два окремих напрямки досліджень, розпочаті в середині-кінці 1800-х років, в кінцевому підсумку призвели до відкриття і характеристики ДНК і основ генетики, науки про спадковість. Ці напрямки досліджень почали сходитися в 1920-х роках, і дослідження з використанням мікробних систем в кінцевому підсумку призвели до значного внеску в з'ясування молекулярної основи генетики.
Відкриття та характеристика ДНК
Сучасне розуміння ДНК еволюціонувало від відкриття нуклеїнової кислоти до розробки моделі подвійної спіралі. У 1860-х роках Фрідріх Мішер (1844—1895), лікар за професією, був першою людиною, яка виділила багаті фосфором хімічні речовини з лейкоцитів (лейкоцитів) з гною на використаних бинтах з місцевої хірургічної клініки. Він назвав ці хімічні речовини (які в кінцевому підсумку були б відомі як РНК і ДНК) «нуклеїнами», оскільки вони були виділені з ядер клітин. Його учень Річард Альтманн (1852—1900) згодом назвав її «нуклеїновою кислотою» через 20 років, коли виявив кислотну природу нуклеїну. В останні два десятиліття 19 століття німецький біохімік Альбрехт Коссель (1853—1927) виділив і охарактеризував п'ять різних нуклеотидних основ, що складають нуклеїнову кислоту. Це аденін, гуанін, цитозин, тимін (в ДНК) і урацил (в РНК). Косселл отримав Нобелівську премію з фізіології або медицини в 1910 році за роботу над нуклеїновими кислотами і за значну роботу над білками, включаючи відкриття гістидину.
Основи генетики
Незважаючи на відкриття ДНК в кінці 1800-х років, асоціації зі спадковістю вчені не робили ще багато десятиліть. Щоб встановити цей зв'язок, вчені, в тому числі ряд мікробіологів, провели безліч експериментів над рослинами, тваринами, бактеріями.
Рослини гороху Менделя
Поки Мішер ізолював та відкривав ДНК у 1860-х роках, австрійський монах і ботанік Йоганн Грегор Мендель (1822—1884) експериментував із садовим горохом, демонструючи та документуючи основні закономірності спадкування, тепер відомі як закони Менделя.
У 1856 році Мендель розпочав свої десятирічні дослідження зразків спадкування. Він використовував диплоїдний садовий горох, Pisum sativum, як свою основну модель системи, оскільки він, природно, самозапліднюється і є дуже інбредним, виробляючи «істинні» лінії рослин гороху - рослини, які завжди дають потомство, схоже на батьків. Експериментуючи з рослинами гороху, Мендель уникав появи несподіваних рис у потомства, які можуть виникнути, якщо він використовував рослини, які не були справжнім розмноженням. Мендель виконував гібридизації, які передбачають спарювання двох істинно селекційних особин (P покоління), що мають різні риси, і досліджував особливості їх потомства (перше синівське покоління, F 1), а також потомство самозапліднення покоління F 1 (друге синівське покоління, F 2) (рис.\(\PageIndex{1}\)).
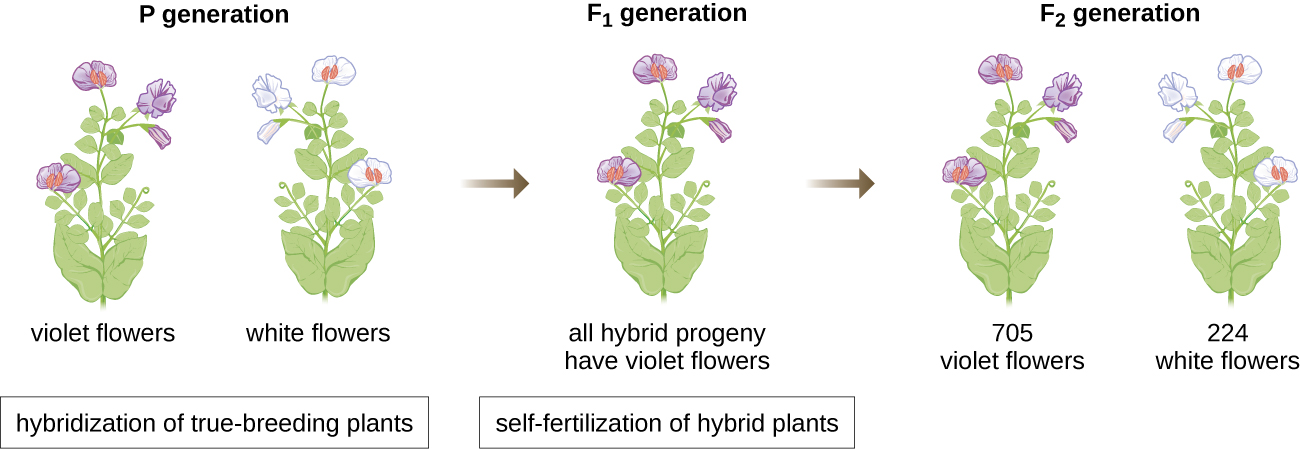
У 1865 році Мендель представив місцевому природничому суспільству результати своїх експериментів з майже 30 000 гороховими рослинами. Він продемонстрував, що риси передаються вірою і правдою від батьків до потомства незалежно від інших рис. У 1866 році він опублікував свою роботу «Експерименти з гібридизації рослин» 1 у Праці товариства природної історії Брунна. Робота Менделя пройшла практично непоміченою науковим співтовариством, яке вірило, неправильно, в теорію змішування рис в безперервних варіаціях.
Він не був визнаний за його надзвичайний науковий внесок протягом життя. Насправді, лише в 1900 році його робота була заново відкрита, відтворена та відновлена вченими на межі виявлення хромосомної основи спадковості.
Хромосомна теорія успадкування
Мендель проводив свої експерименти задовго до того, як хромосоми були візуалізовані під мікроскопом. Однак з удосконаленням мікроскопічних методик в кінці 1800-х років клітинні біологи могли фарбувати і візуалізувати субклітинні структури барвниками і спостерігати за їх діями під час мейозу. Вони змогли спостерігати хромосоми, що розмножуються, конденсуються з аморфної ядерної маси в різні Х-подібні тіла і мігрують до окремих клітинних полюсів. Припущення про те, що хромосоми можуть бути ключем до розуміння спадковості, змусили кількох вчених вивчити публікації Менделя та переоцінити його модель з точки зору поведінки хромосом під час мітозу та мейозу.
У 1902 році Теодор Бовері (1862—1915) спостерігав, що у морських їжаків ядерні компоненти (хромосоми) визначали правильний ембріональний розвиток. Того ж року Уолтер Саттон (1877—1916) спостерігав поділ хромосом на дочірні клітини під час мейозу. Разом ці спостереження привели до розвитку хромосомної теорії успадкування, яка ідентифікувала хромосоми як генетичний матеріал, відповідальний за успадкування Менделя.
Незважаючи на переконливі кореляції між поведінкою хромосом під час мейозу і спостереженнями Менделя, хромосомна теорія успадкування була запропонована задовго до того, як з'явилися будь-які прямі докази того, що риси були перенесені на хромосоми. Томас Хант Морган (1866—1945) і його колеги провели кілька років, виконуючи схрещування з дрозофілою меланогастер. Вони виконували ретельні мікроскопічні спостереження хромосом мух і співвіднесли ці спостереження з результуючими характеристиками мух. Їх робота надала перші експериментальні докази, що підтверджують хромосомну теорію успадкування на початку 1900-х років. У 1915 році Морган і його колеги по «Fly Room» опублікували Механізм менделівської спадковості, який ідентифікував хромосоми як клітинні структури, відповідальні за спадковість. За свій значний внесок у генетику Морган отримав Нобелівську премію з фізіології або медицини в 1933 році.
Наприкінці 1920-х років Барбара МакКлінток (1902—1992) розробила методи хромосомного фарбування для візуалізації та диференціації різних хромосом кукурудзи (кукурудзи). У 1940-х і 1950-х роках вона визначила подію розриву хромосоми 9, яку вона назвала локусом дисоціації (Ds). Ds міг змінювати положення всередині хромосоми. Вона також визначила локус активатора (Ac). Розрив хромосоми Ds може бути активований елементом Ac (ферментом транспозази). Спочатку знаходження МакКлінтоком цих стрибаючих генів, які ми зараз називаємо транспозонами, наукове співтовариство не прийняло. Лише в 1960-х роках і пізніше транспозони були виявлені в бактеріофагах, бактеріях та дрозофілах. Сьогодні ми знаємо, що транспозони - це рухливі сегменти ДНК, які можуть переміщатися всередині генома організму. Вони можуть регулювати експресію генів, експресію білка та вірулентність (здатність викликати захворювання).
Мікроби та віруси в генетичних дослідженнях
Мікробіологи також відіграли вирішальну роль у нашому розумінні генетики. Експериментальні організми, такі як садовий горох Менделя, плодові мухи Моргана та кукурудза МакКлінтока, вже успішно використовувались, щоб прокласти шлях до розуміння генетики. Однак мікроби та віруси були (і досі є) чудовими модельними системами для вивчення генетики, оскільки, на відміну від гороху, дрозофіл та кукурудзи, вони легше розмножуються в лабораторії, зростаючи до високої щільності населення на невеликій кількості простору та за короткий час. Крім того, через їх структурної простоти мікроби і віруси легше маніпулюють генетично.
На щастя, незважаючи на значні відмінності в розмірах, структурі, стратегіях розмноження та інших біологічних характеристиках, існує біохімічна єдність між усіма організмами; вони мають спільні ті ж основні молекули, відповідальні за спадковість та використання генетичного матеріалу для надання клітинам їх варіативних характеристики. За словами французького вченого Жака Монода, «Те, що вірно для кишкової палички, вірно і для слона», що означає, що біохімія життя підтримувалася протягом еволюції і поділяється у всіх формах життя, від простих одноклітинних організмів до великих складних організмів. Ця біохімічна безперервність робить мікроби чудовими моделями для використання для генетичних досліджень.
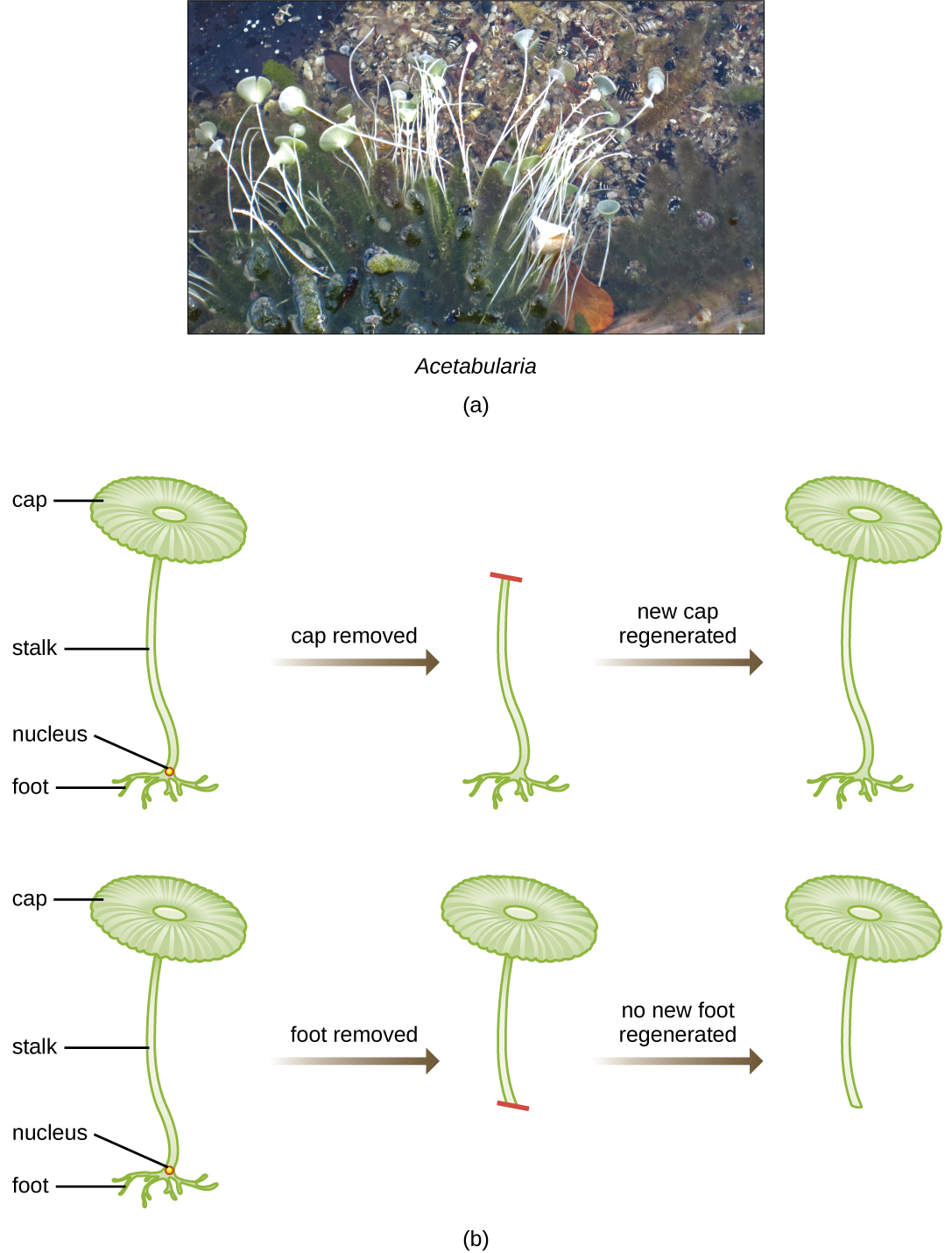
У розумному наборі експериментів у 1930-х і 1940-х роках німецький вчений Йоахім Геммерлінг (1901—1980), використовуючи одноклітинну водорость Acetabularia як мікробну модель, встановив, що генетична інформація в еукаріотичній клітині розміщується в ядрі. Acetabularia spp. - це надзвичайно великі водорості клітини, які ростуть асиметрично, утворюючи «стопу», що містить ядро, яке використовується для кріплення субстрату; стебло; і парасолькоподібний ковпачок - структури, які все можна легко побачити неозброєним оком. У ранньому наборі експериментів Hämmerling видалив або шапку, або ніжку клітин і спостерігав, чи нові шапки або ноги були регенеровані (рис.\(\PageIndex{2}\)). Він виявив, що при видаленні стопи цих клітин нові ноги не ростуть; однак, коли з клітин знімали ковпачки, нові ковпачки регенерували. Це припускало, що спадкова інформація була розташована в ядросодержащей ніжці кожної клітини.
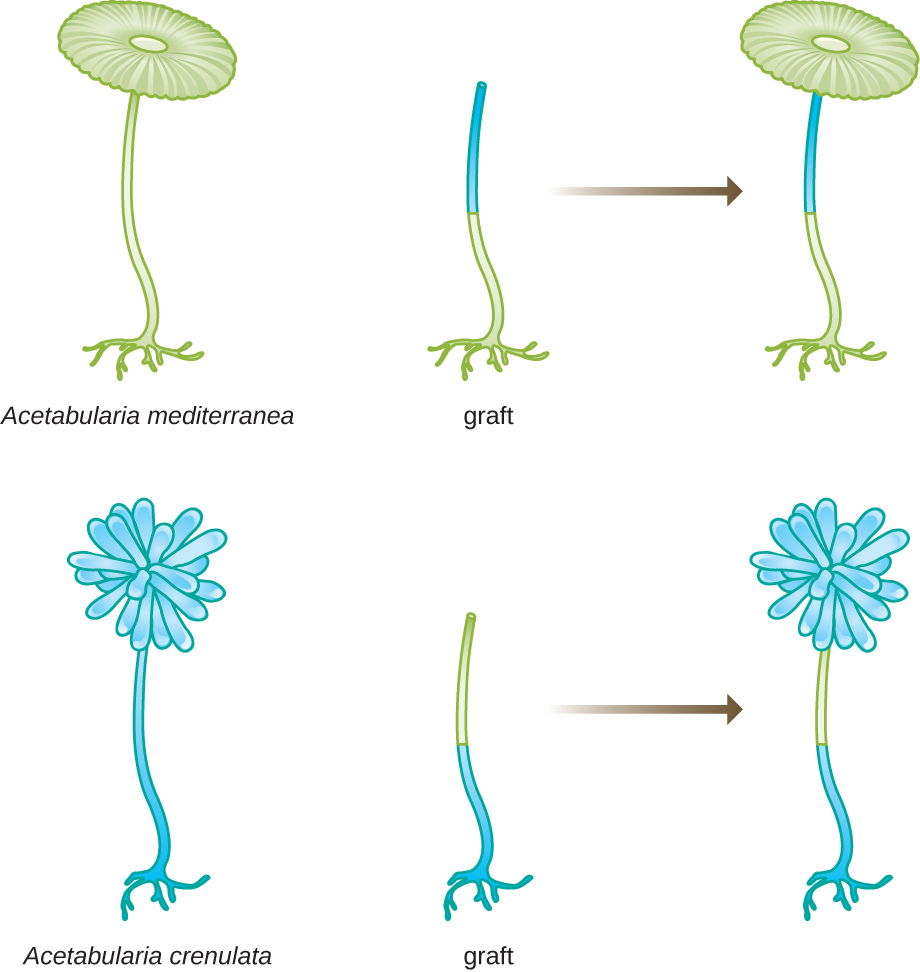
В іншому наборі експериментів Hämmerling використовував два види Acetabularia, які мають різну морфологію капелюшка, A. crenulata і A. mediterranea (рис.\(\PageIndex{3}\)). Він вирізав капелюшки з обох типів клітин, а потім прищепив стебло з A. crenulata на стопу A. mediterranea, і навпаки. Згодом він помітив, що щеплена клітина з стопою A. crenulata і стеблом A. mediterranea розвинула капелюшок з морфологією A. crenulata. І навпаки, щеплена клітина з стопою A. mediterranea та стеблом A. crenulata розвинула капелюшок з морфологією A. mediterranea. Він мікроскопічно підтвердив наявність ядер в стопах цих клітин і приписував розвиток цих морфологій капелюшків ядру кожної щепленої клітини. Таким чином, він експериментально показав, що ядро - це місце розташування генетичного матеріалу, який диктував властивості клітини.
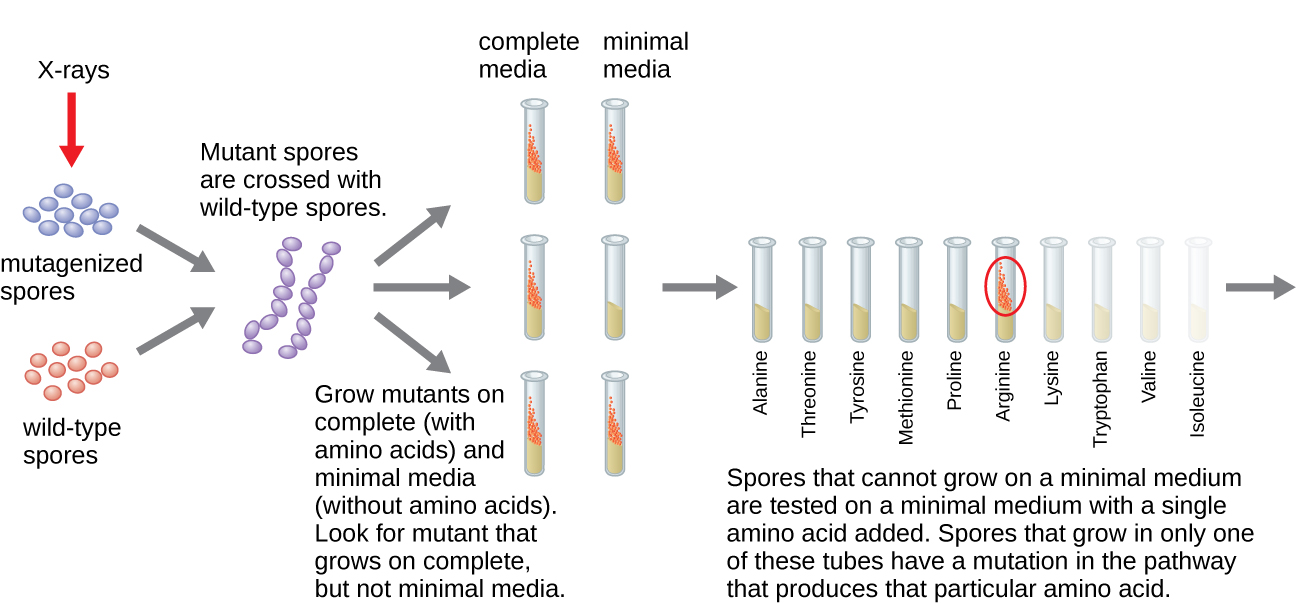
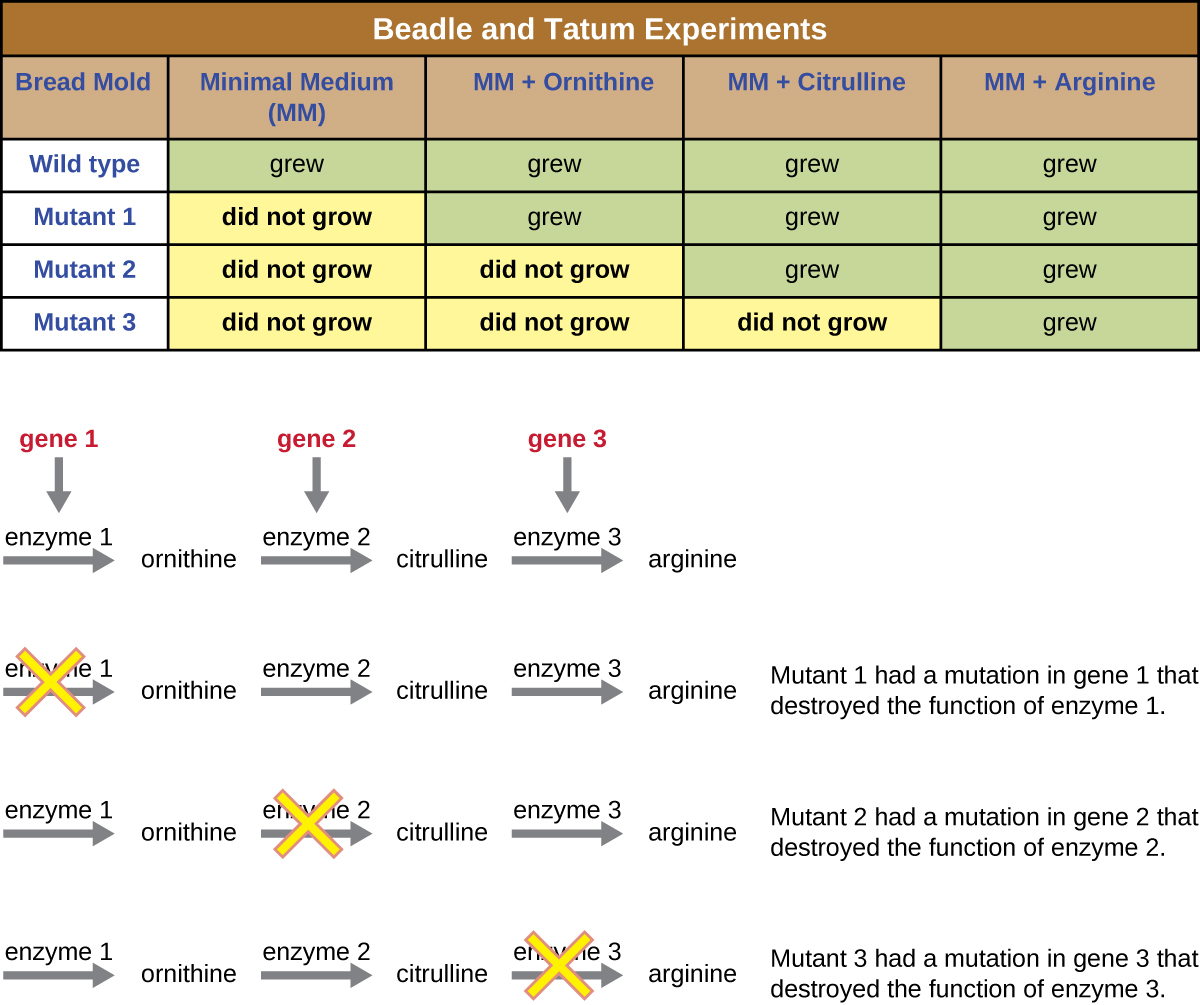
Іншу мікробну модель, червону хлібну цвіль Neurospora crassa, використовували Джордж Бідл та Едвард Татум, щоб продемонструвати взаємозв'язок між генами та білками, які вони кодують. Бідл працював з фруктовими мухами в лабораторії Моргана, але виявив їх занадто складними для виконання певних типів експериментів. N. crassa, з іншого боку, є простішим організмом і має здатність рости на мінімальному середовищі, оскільки містить ферментативні шляхи, які дозволяють йому використовувати середовище для виробництва власних вітамінів та амінокислот.
Бідл і Татум опромінювали цвіль рентгенівськими променями, щоб викликати зміни послідовності нуклеїнових кислот, які називаються мутаціями. Вони спаровували опромінені спори цвілі і намагалися виростити їх як на повному середовищі, так і на мінімальному середовищі. Вони шукали мутантів, які росли на повноцінному середовищі, доповнювалися вітамінами і амінокислотами, але не росли на мінімальному середовищі, де бракує цих добавок. Такі цвілі теоретично містили мутації в генах, які кодували біосинтетичні шляхи. Виявивши таких мутантів, вони систематично тестували кожного, щоб визначити, який вітамін або амінокислоту він не зміг виробити (рис.\(\PageIndex{4}\)) і опублікували цю роботу в 1941 році.
Подальша робота Бідл, Татума та колег показала, що вони можуть виділити різні класи мутантів, які вимагали певної добавки, як амінокислота аргінін (рис.\(\PageIndex{5}\)). Маючи певні знання про шлях біосинтезу аргініну, вони визначили три класи мутантів аргініну, доповнюючи мінімальне середовище проміжними продуктами (цитрулін або орнітин) у шляху. Три мутанти відрізнялися своєю здатністю рости в кожному із засобів масової інформації, що змусило групу вчених у 1945 році запропонувати, що кожен тип мутантів мав дефект іншого гена в шляху біосинтезу аргініну. Це призвело до так званої гіпотези про один ген-один фермент, яка припускала, що кожен ген кодує один фермент.
Подальші знання про процеси транскрипції та перекладу змусили вчених переглянути це до гіпотези «один ген—один поліпептид». Хоча є деякі гени, які не кодують поліпептиди (а скоріше кодують для трансферних РНК [тРНК] або рибосомних РНК [РРНК], про які ми поговоримо пізніше), гіпотеза про один ген-один фермент вірна у багатьох випадках, особливо у мікробів. Відкриття Бідлом і Татумом зв'язку між генами і відповідними характеристиками принесло їм Нобелівську премію з фізіології та медицини 1958 року і з тих пір стало основою сучасної молекулярної генетики.
Щоб дізнатися більше про експерименти Beadle і Tatum, відвідайте цей веб-сайт з навчального центру ДНК.
Вправа\(\PageIndex{2}\)
- Який організм використовували Морган і його колеги для розробки хромосомної теорії успадкування? Які риси вони відстежували?
- Що довев Хеммерлінг своїми експериментами над ацетабулярією?
ДНК як молекула, відповідальна за спадковість
До початку 20 століття вже була зроблена велика робота над характеристикою ДНК та встановленням основ генетики, включаючи приписування спадковості хромосомам, знайденим всередині ядра. Незважаючи на всі ці дослідження, лише в 20 столітті ці напрямки досліджень зійшлися, і вчені почали вважати, що ДНК може бути генетичним матеріалом, який потомство успадкувало від батьків. Вважалося, що ДНК, що містить лише чотири різних нуклеотиди, структурно занадто проста для кодування такої складної генетичної інформації. Замість цього вважалося, що білок має складність, необхідну для роботи в якості клітинної генетичної інформації, оскільки він складається з 20 різних амінокислот, які можуть бути об'єднані у величезній кількості комбінацій. Мікробіологи зіграли ключову роль у дослідженні, які визначили, що ДНК - це молекула, відповідальна за спадковість.
Експерименти трансформації Гріффіта
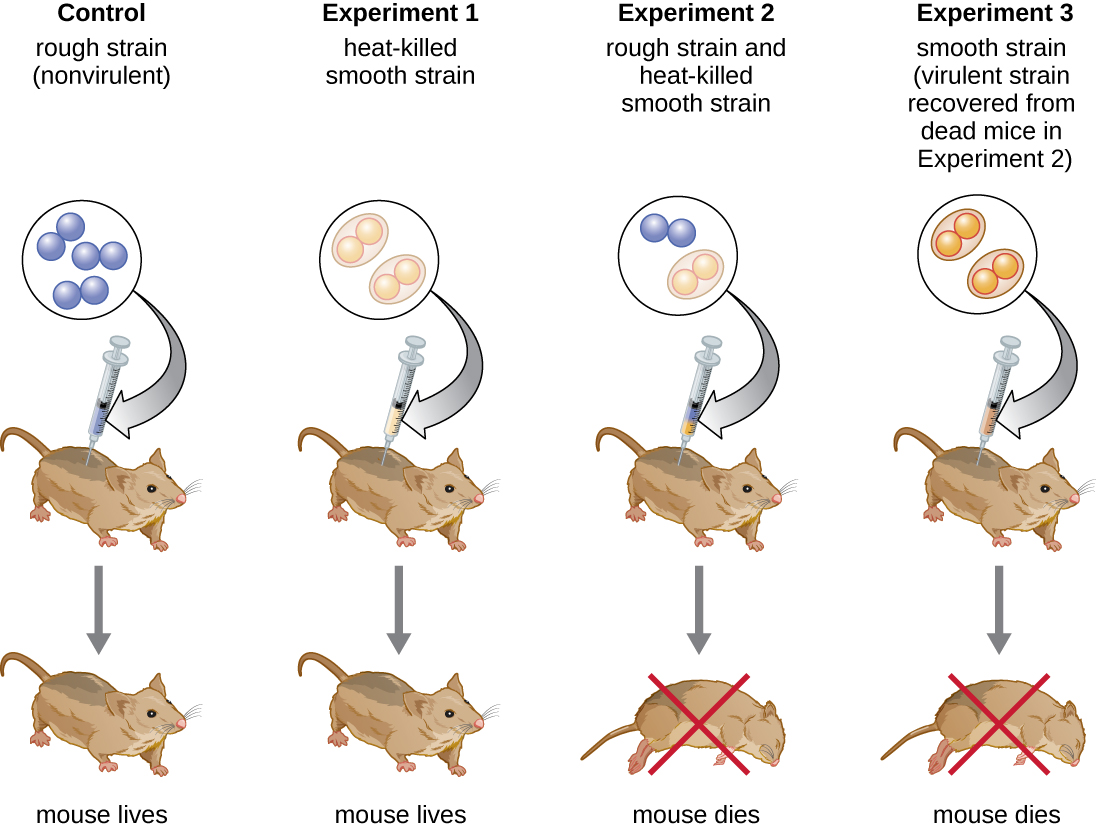
Британський бактеріолог Фредерік Гріффіт (1879—1941) був, мабуть, першою людиною, яка показала, що спадкова інформація може передаватися з однієї клітини в іншу «горизонтально» (між членами одного покоління), а не «вертикально» (від батька до потомства). У 1928 році він повідомив про першу демонстрацію бактеріальної трансформації - процесу, при якому зовнішня ДНК захоплюється клітиною, тим самим змінюючи її характеристики. 3 Він працював з двома штамами Streptococcus pneumoniae, бактерією, яка викликає пневмонію: грубим (R) штамом та гладким (S) штамом. Штам R є непатогенним і не має капсули на зовнішній поверхні; в результаті колонії з штаму R при вирощуванні на пластинках виявляються шорсткими. Штам S є патогенним і має капсулу поза клітинною стінкою, що дозволяє йому уникнути фагоцитозу імунною системою господаря. Капсули призводять до того, що колонії з штаму S здаються гладкими при вирощуванні на тарілках.
У серії експериментів Гріффіт проаналізував вплив живих R, живих S та термоубитих S штамів S. pneumoniae на живих мишей (рис.\(\PageIndex{6}\)). Коли мишам вводили живий штам S, миші гинули. Коли він вводив мишам живий штам R або штам S, знищений теплом, миші вижили. Але коли він вводив мишам суміш живого штаму R і теплового штаму S, миші загинули. Виділивши живі бактерії від мертвої миші, він лише відновив S штам бактерій. Коли він потім вводив цей ізольований штам S свіжим мишам, миші загинули. Гріффіт дійшов висновку, що щось перейшло від нагрітого штаму S в живий штам R і «перетворило» його в патогенний штам S; він назвав це «принципом трансформації». Ці експерименти тепер відомі як експерименти трансформації Гріффіта.
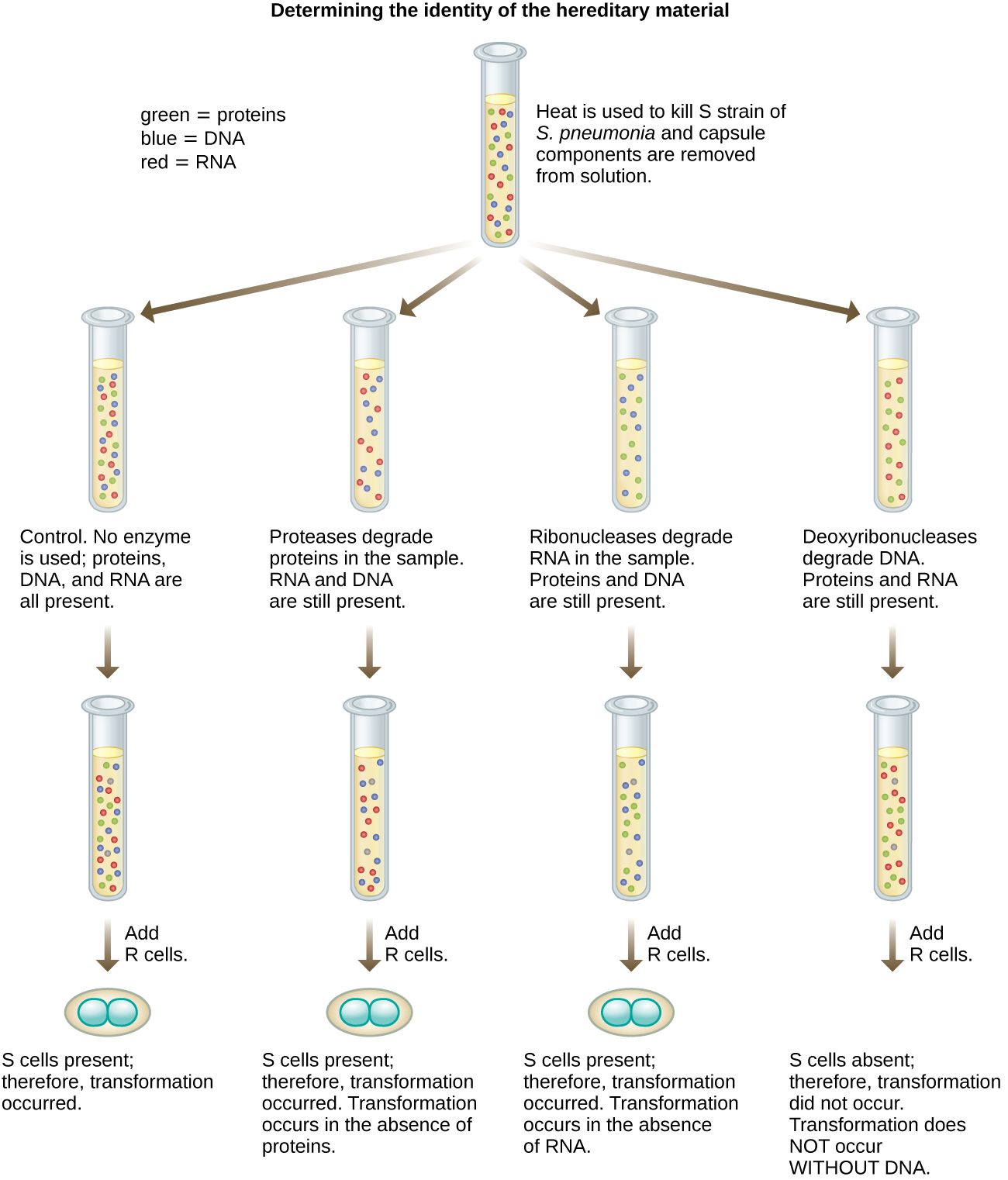
У 1944 році Освальд Ейвері, Колін Маклауд і Маклін Маккарті були зацікавлені в подальшому вивченні принципу трансформації Гріффіта. Вони виділили штам S від заражених мертвих мишей, тепло вбили його, і інактивували різні компоненти екстракту S, проводячи систематичне елімінаційне дослідження (рис.\(\PageIndex{7}\)). Вони використовували ферменти, які спеціально розщеплювали білки, РНК та ДНК, і змішували екстракт S з кожним із цих індивідуальних ферментів. Потім вони перевірили результуючу здатність кожної комбінації екстракту/ферменту перетворювати штам R, що спостерігається дифузним ростом штаму S в культуральних середовищах і підтверджено візуально ростом на пластинках. Вони виявили, що при деградації ДНК отримана суміш більше не здатна трансформувати бактерії штаму R, тоді як жодна інша ферментативна обробка не змогла запобігти трансформації. Це призвело їх до висновку, що ДНК є трансформаційним принципом. Незважаючи на їх результати, багато вчених не прийняли їх висновок, натомість вважаючи, що в їх екстрактах є білкові забруднювачі.
Вправа\(\PageIndex{3}\)
Як експерименти Ейвері, Маклауда та Маккарті показали, що ДНК була принципом трансформації, вперше описаним Гріффітом?
Доказ ДНК Герші та Чейза як генетичного матеріалу
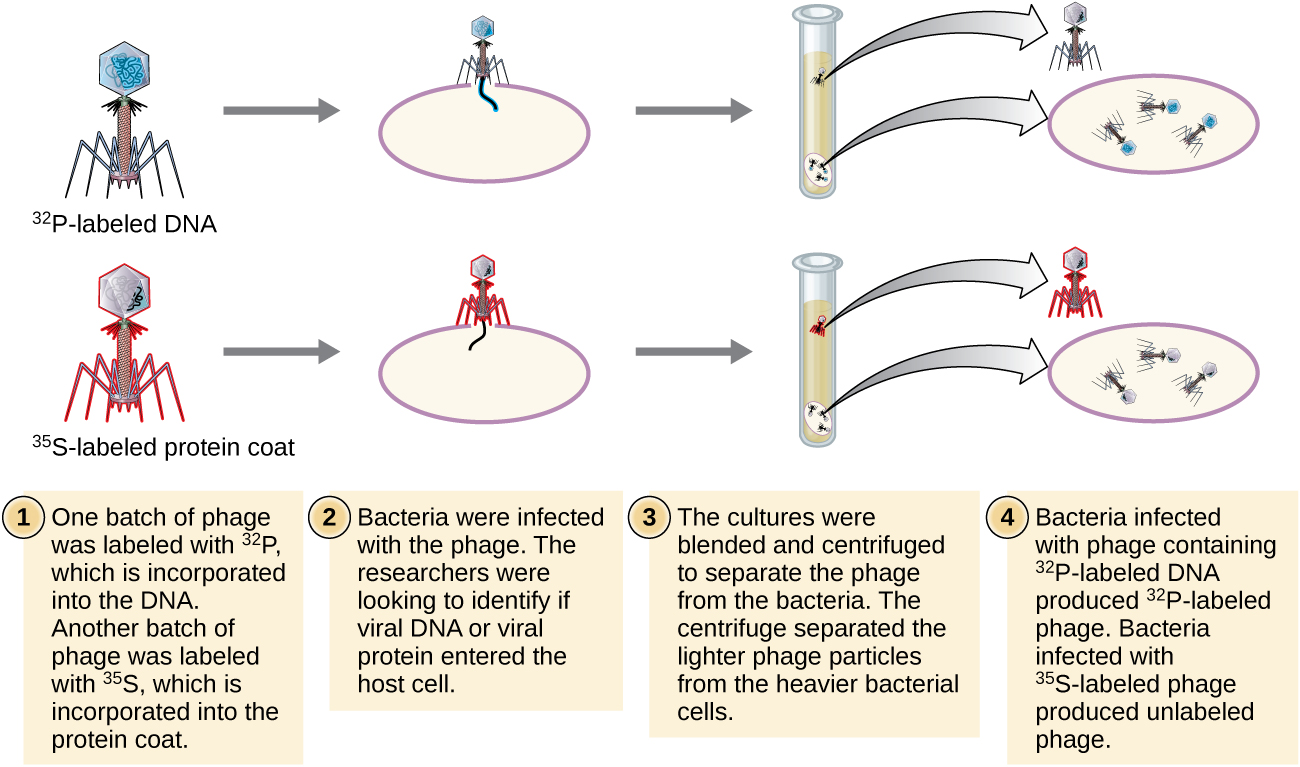
Альфред Херші і Марта Чейз провели власні експерименти в 1952 році і змогли надати підтверджуючі докази того, що генетичним матеріалом була ДНК, а не білок (рис.\(\PageIndex{8}\)). 4 Герші і Чейз вивчали бактеріофаг, вірус, який заражає бактерії. Віруси зазвичай мають просту структуру: білкову оболонку, звану капсидом, і ядро нуклеїнової кислоти, яке містить генетичний матеріал, або ДНК, або РНК (див. Віруси). Особливим бактеріофагом, який вони вивчали, був бактеріофаг Т2, який заражає клітини кишкової палички. Як ми зараз знаємо сьогодні, Т2 прикріплюється до поверхні бактеріальної клітини, а потім він впорскує свої нуклеїнові кислоти всередину клітини. ДНК фагів робить кілька копій себе за допомогою механізми-господаря, і в кінцевому підсумку клітина господаря лопається, виділяючи велику кількість бактеріофагів.
Герші і Чейз позначили білкову оболонку в одній партії фагів, використовуючи радіоактивну сірку, 35 S, оскільки сірка міститься в амінокислотах метіонін і цистеїн, але не в нуклеїнових кислотах. Вони позначили ДНК в іншій партії, використовуючи радіоактивний фосфор, 32 P, оскільки фосфор міститься в ДНК та РНК, але зазвичай не в білку.
Кожній партії фага дозволялося інфікувати клітини окремо. Після зараження Херші і Чейз кладуть кожну фагову бактеріальну суспензію в блендер, який від'єднував оболонки фагів від клітини-господаря, і закручують отриману суспензію в центрифугу. Більш важкі бактеріальні клітини осіли і утворили гранулу, тоді як більш легкі частинки фагів залишилися в супернатанті. У пробірці з позначеним білком радіоактивність залишилася тільки у супернатанта. У трубці з маркуванням ДНК радіоактивність виявлялася тільки в бактеріальних клітині. Герші і Чейз прийшли до висновку, що саме фагова ДНК була введена в клітину, яка несла інформацію для отримання більшої кількості частинок фагів, тим самим доводячи, що джерелом генетичного матеріалу є ДНК, а не білки. В результаті їх роботи наукове співтовариство більш широко прийняло ДНК як молекулу, що відповідає за спадковість.
На той час, як Герші і Чейз опублікували свій експеримент на початку 1950-х років, мікробіологи та інші вчені досліджували спадковість понад 80 років. Спираючись на дослідження один одного в той час, завершився загальною згодою про те, що ДНК є генетичним матеріалом, відповідальним за спадковість (рис.\(\PageIndex{9}\)). Ці знання створили основу для настання епохи молекулярної біології та значних досягнень у біотехнології та системній біології, які ми переживаємо сьогодні.
Щоб дізнатися більше про експерименти, пов'язані з історією генетики та відкриттям ДНК як генетичного матеріалу клітин, відвідайте цей веб-сайт з навчального центру ДНК.
Вправа\(\PageIndex{4}\)
Як Герші і Чейз використовували мікроби, щоб довести, що ДНК є генетичним матеріалом?
Ключові поняття та резюме
- ДНК була виявлена і охарактеризована задовго до того, як була зрозуміла її роль у спадковості. Мікробіологи зіграли значну роль у демонстрації того, що ДНК - це спадкова інформація, знайдена всередині клітин.
- У 1850-х і 1860-х роках Грегор Мендель експериментував з істинним садовим горохом, щоб продемонструвати спадковість конкретних спостережуваних ознак.
- У 1869 році Фрідріх Мішер виділив і очистив з ядер лейкоцитів з'єднання, багате фосфором; він назвав з'єднання нуклеїном. Учень Мішера Річард Альтманн відкрив його кислотну природу, перейменуючи його нуклеїновою кислотою. Альбрехт Косселл охарактеризував нуклеотидні основи, знайдені в нуклеїнових кислотах.
- Хоча Уолтер Саттон і Теодор Бовері запропонували хромосомну теорію спадкування в 1902 році, вона не була науково продемонстрована до публікації 1915 року роботи Томаса Ханта Моргана та його колег.
- Використовуючи ацетабулярію, велику водорослеву клітину, як свою модельну систему, Йоахім Геммерлінг продемонстрував у 1930-х і 1940-х роках, що ядро є місцем розташування спадкової інформації в цих клітині.
- У 1940-х роках Джордж Бідл та Едвард Татум використовували цвіль Neurospora crassa, щоб показати, що виробництво кожного білка знаходиться під контролем одного гена, демонструючи гіпотезу «один ген—один фермент».
- У 1928 році Фредерік Гріффіт показав, що мертві інкапсульовані бактерії можуть передавати генетичну інформацію жити некапсульованим бактеріям і перетворювати їх на шкідливі штами. У 1944 році Освальд Ейвері, Колін Маклауд і Маклін Маккарті визначили з'єднання як ДНК.
- Природа ДНК як молекули, яка зберігає генетичну інформацію, була однозначно продемонстрована в експерименті Альфреда Херші і Марти Чейз, опублікованому в 1952 році. Мічені ДНК з бактеріальних вірусів проникли і інфіковані бактеріальні клітини, породжуючи більше вірусних частинок. Марковані білкові оболонки не брали участі в передачі генетичної інформації.
Виноски
- Мендель Ю.Г. «Версуше убер Pflanzenhybriden». Видалення природних ресурсів Vereines в Брюнн, Bd. Припинення 4 (1865) :3—7. (Для перекладу англійською дивіться http://www.mendelweb.org/Mendel.plain.html)
- Бідл Г.В., Татум Е.Л. «Генетичний контроль біохімічних реакцій в нейроспорі». Вісник Національної академії наук 27 № 11 (1941) :499—506.
- 3 Ф.Гріффіт. «Значення пневмококових типів». Журнал гігієни 27 № 2 (1928) :8—159.
- 4 А.Д. Херші, М.Чейз. «Незалежні функції вірусного білка та нуклеїнової кислоти у рості бактеріофагів». Журнал загальної фізіології 36 № 1 (1952) :39—56.