8.6: Фотосинтез та важливість світла
- Page ID
- 3938

Цілі навчання
- Опишіть функцію та розташування фотосинтетичних пігментів у еукаріотів та прокаріотів
- Охарактеризуйте основні продукти світлозалежних і світлонезалежних реакцій
- Опишіть реакції, які виробляють глюкозу в фотосинтетичній клітині
- Порівняйте і контрастуйте циклічне і нециклічне фотофосфорилювання
Гетеротрофні організми, починаючи від кишкової палички до людини, покладаються на хімічну енергію, що міститься в основному в молекулах вуглеводів. Багато з цих вуглеводів виробляються фотосинтезом, біохімічним процесом, за допомогою якого фототрофні організми перетворюють сонячну енергію (сонячне світло) в хімічну енергію. Хоча фотосинтез найчастіше асоціюється з рослинами, мікробний фотосинтез також є значним постачальником хімічної енергії, підживлюючи багато різноманітних екосистем. У цьому розділі мова піде про мікробний фотосинтез.
Фотосинтез відбувається в дві послідовні стадії: світлозалежні реакції і світлонезалежні реакції (рис.\(\PageIndex{1}\)). У світлозалежних реакціях енергія сонячного світла поглинається молекулами пігменту в фотосинтетичних мембранах і перетворюється в накопичену хімічну енергію. У незалежних від світла реакціях хімічний енер, що виробляється світлозалежними реакціями, використовується для керування збіркою молекул цукру за допомогою CO 2; однак ці реакції все ще легкозалежні, оскільки продукти світлозалежних реакцій, необхідних для їх керування, короткі -жив. Світлозалежні реакції виробляють АТФ і або NADPH, або NADH для тимчасового зберігання енергії. Ці енергоносії використовуються в світлонезалежних реакціях для приводу енергетично несприятливого процесу «фіксації» неорганічного СО 2 в органічній формі, цукру.
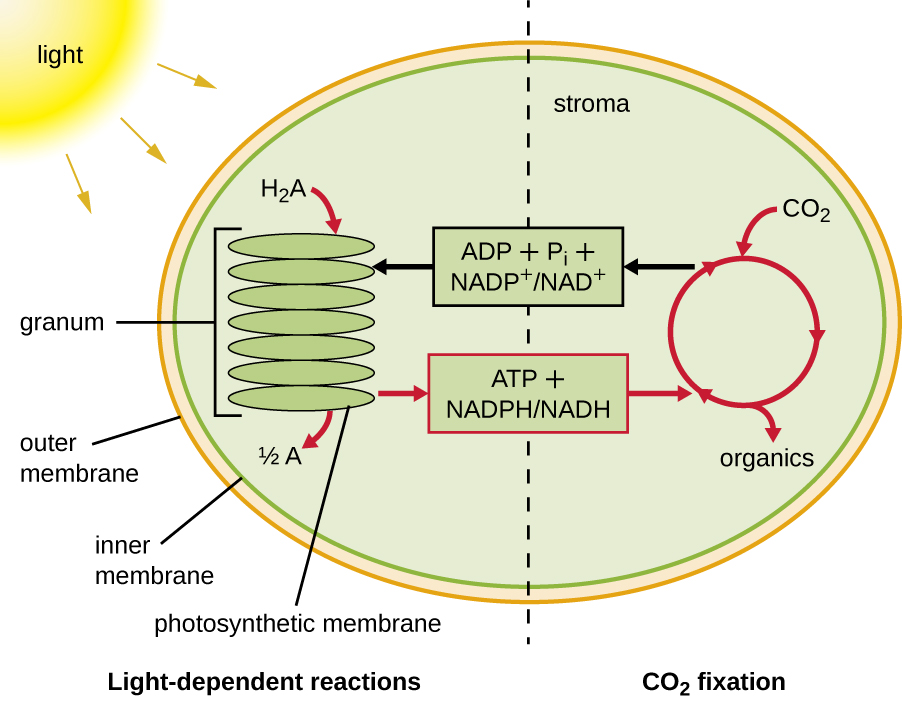
Фотосинтетичні структури в еукаріотів і прокаріотів
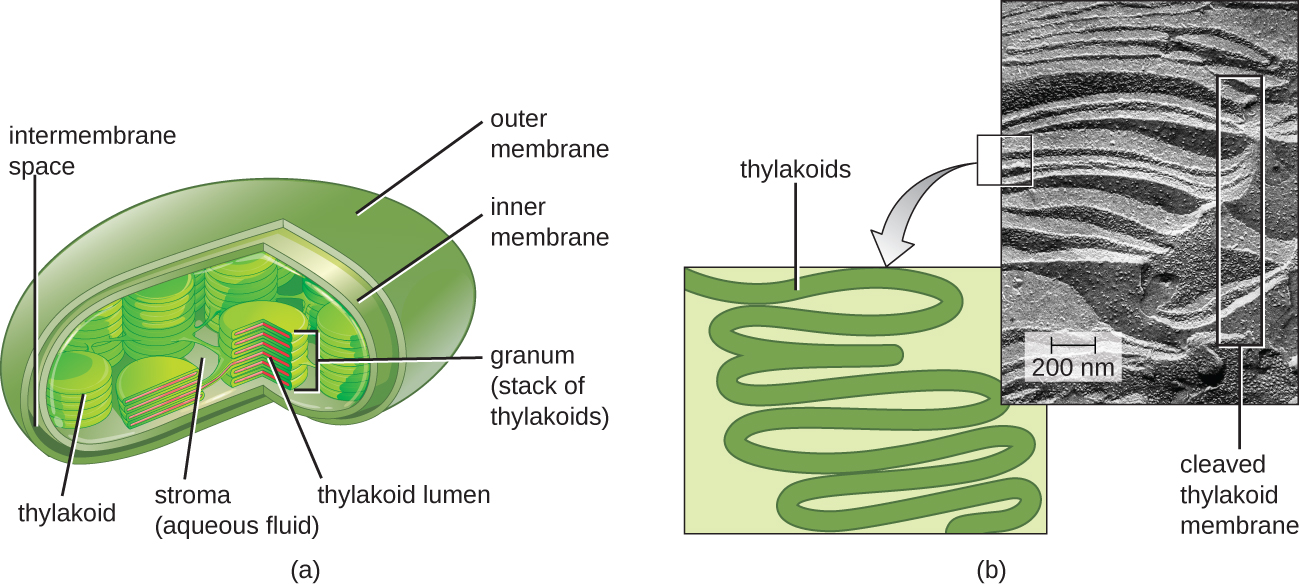
У всіх фототрофних еукаріотів фотосинтез відбувається всередині хлоропласту, органели, яка виникла у еукаріотів шляхом ендосимбіозу фотосинтетичної бактерії (див. Унікальні характеристики еукаріотичних клітин). Ці хлоропласти укладені подвійною мембраною з внутрішнім і зовнішнім шарами. Усередині хлоропласту знаходиться третя мембрана, яка утворює складені, дископодібні фотосинтетичні структури, звані тилакоїдами (рис.\(\PageIndex{2}\)). Стек тилакоїдів називається гранумом, а простір, що оточує грануму всередині хлоропласту, називається стромою.
Фотосинтетичні мембрани у прокаріотів, навпаки, не організовані в окремі мембранно-закриті органели; скоріше, вони являють собою складені ділянки плазматичної мембрани. Наприклад, у ціанобактерій ці складені області також називають тилакоїдами. У будь-якому випадку вбудовані в тилакоїдні мембрани або інші фотосинтетичні бактеріальні мембрани є фотосинтетичними молекулами пігменту, організованими в одну або кілька фотосистем, де світлова енергія фактично перетворюється в хімічну енергію.
Фотосинтетичні пігменти всередині фотосинтетичних мембран організовані в фотосистеми, кожна з яких складається з світлозбирального (вусикового) комплексу і реакційного центру. Світлозбиральний комплекс складається з безлічі білків і пов'язаних з ними пігментів, які кожен може поглинати світлову енергію і, таким чином, збуджуватися. Ця енергія передається від однієї молекули пігменту до іншої, поки в кінцевому підсумку (приблизно через мільйонну частку секунди) вона не буде доставлена в реакційний центр. До цього моменту між молекулами передавалася лише енергія, а не електрони. Реакційний центр містить молекулу пігменту, яка може піддаватися окисленню при збудженні, фактично віддаючи електрон. Саме на цьому етапі фотосинтезу світлова енергія перетворюється в збуджений електрон.
Різні види світлозбиральних пігментів поглинають унікальні візерунки довжин хвиль (кольорів) видимого світла. Пігменти відображають або передають довжини хвиль, які вони не можуть поглинати, завдяки чому вони виглядають відповідним кольором. Прикладами фотосинтетичних пігментів (молекул, що використовуються для поглинання сонячної енергії) є бактеріохлорофіли (зелений, фіолетовий або червоний), каротиноїди (помаранчевий, червоний або жовтий), хлорофіли (зелений), фікоціаніни (синій) та фікоеритрини (червоний). Маючи суміші пігментів, організм може поглинати енергію з більшої довжини хвиль. Оскільки фотосинтетичні бактерії зазвичай ростуть у конкуренції за сонячне світло, кожен тип фотосинтетичних бактерій оптимізований для збору довжин хвиль світла, яким він зазвичай піддається, що призводить до розшарування мікробних спільнот у водних та ґрунтових екосистемах за якістю світла та проникненням.
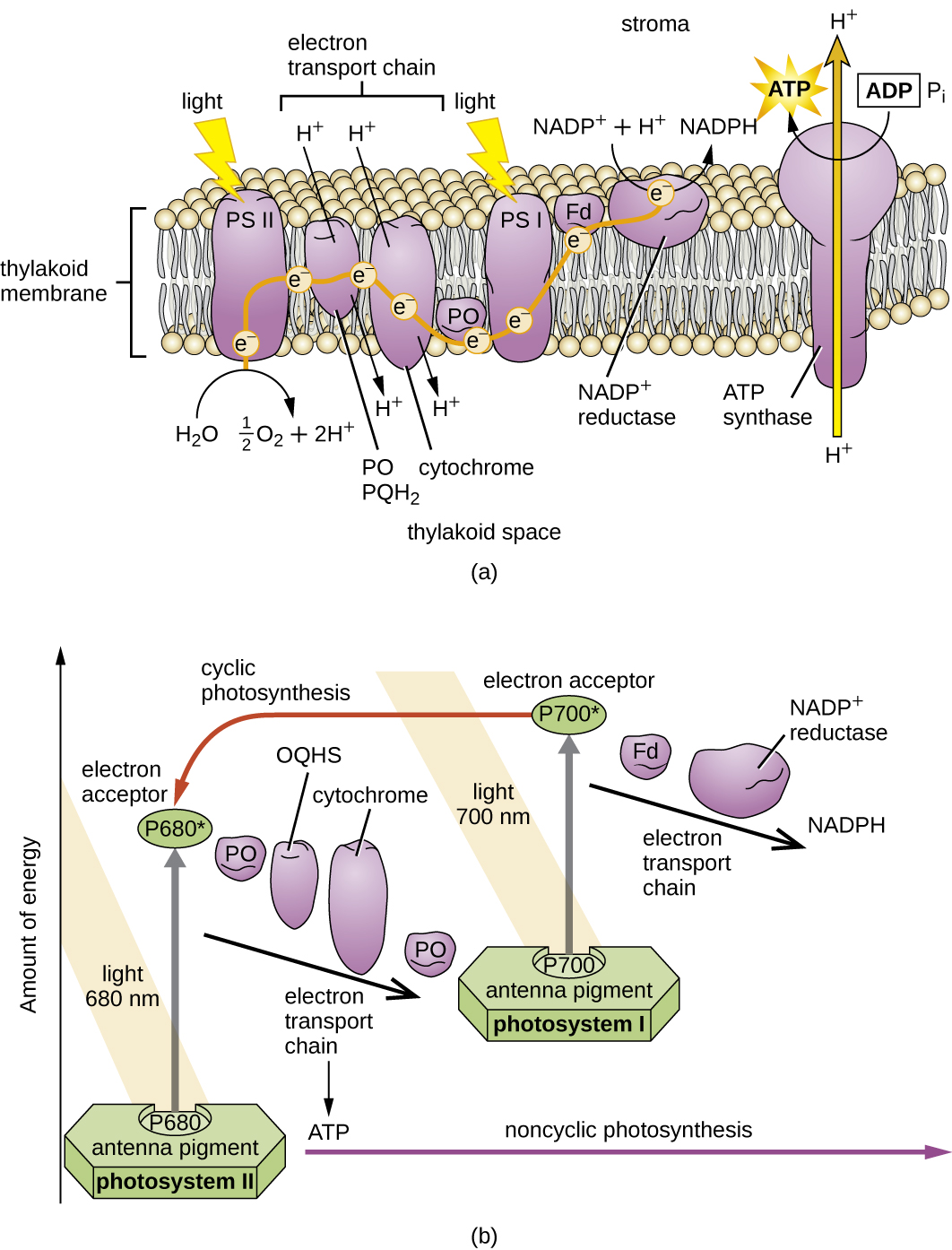
Після того, як комплекс збору світла передає енергію в реакційний центр, реакційний центр доставляє свої високоенергетичні електрони один за іншим до носія електронів в транспортній системі електронів, і ініціюється перенесення електронів через ETS. ETS схожий на той, що використовується в клітинному диханні і вбудований в фотосинтетичну мембрану. Зрештою, електрон використовується для отримання NADH або NADPH. Електрохімічний градієнт, який утворюється через фотосинтетичну мембрану, використовується для генерації АТФ шляхом хеміосмосу через процес фотофосфорилювання, ще один приклад окислювального фосфорилювання (рис.\(\PageIndex{3}\)).
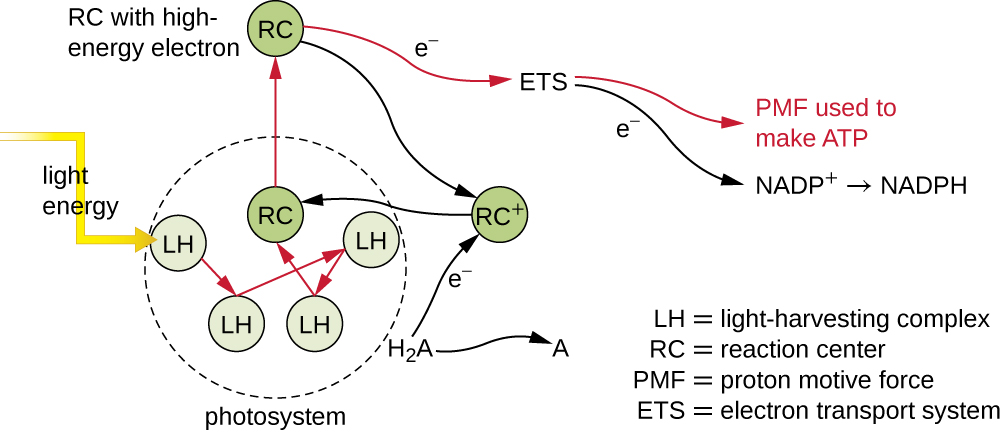
Вправа\(\PageIndex{1}\)
У фототрофного еукаріот, де відбувається фотосинтез?
Кисневий і аноксигенний фотосинтез
Щоб фотосинтез продовжувався, електрон, втрачений з пігменту реакційного центру, повинен бути замінений. Джерело цього електрона (Н 2 А) диференціює кисневий фотосинтез рослин і ціанобактерій від аноксигенного фотосинтезу, здійсненого іншими видами фототрофів бактерій (рис.\(\PageIndex{4}\)). При кисневому фотосинтезі Н 2 О розщеплюється і постачає електрон в реакційний центр. Оскільки кисень генерується як побічний продукт і вивільняється, цей тип фотосинтезу називають кисневим фотосинтезом. Однак, коли інші відновлені сполуки служать донором електронів, кисень не утворюється; ці типи фотосинтезу називаються аноксигенним фотосинтезом. Донором електронів\(\ce{(S2O3^{2-})}\) можуть служити сірководень (H 2 S) або тіосульфат, утворюючи в результаті елементарні\(\ce{(SO4^{2-})}\) іони сірки і сульфату відповідно.
Фотосистеми були класифіковані на два типи: фотосистема I (PSI) і фотосистема II (PSII) (рис.\(\PageIndex{5}\)). Ціанобактерії та рослинні хлоропласти мають обидві фотосистеми, тоді як аноксигенні фотосинтетичні бактерії використовують лише одну з фотосистем. Обидві фотосистеми збуджуються світловою енергією одночасно. Якщо клітина вимагає і АТФ, і НАДФГ для біосинтезу, то вона буде здійснювати нециклічне фотофосфорилювання. При передачі електрона реакційного центру PSII до ЕТС, що з'єднує PSII і PSI, втрачений електрон з реакційного центру PSII замінюється розщепленням води. Збуджений електрон реакційного центру PSI використовується для зменшення NADP + до NADPH і замінюється електроном, що виходить з ETS. Потік електронів таким способом називається Z-схемою.
Якщо потреба клітини в АТФ значно більша, ніж її потреба в NADPH, вона може обійти виробництво відновлювальної потужності за допомогою циклічного фотофосфорилювання. Тільки PSI використовується під час циклічного фотофосфорилювання; високоенергетичний електрон реакційного центру PSI передається носію ETS, а потім в кінцевому підсумку повертається до окисленого пігменту реакційного центру PSI, тим самим зменшуючи його.
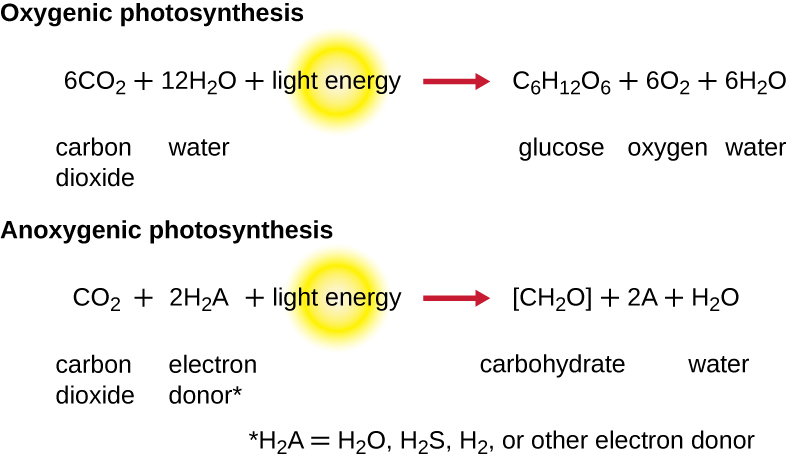
Вправа\(\PageIndex{2}\)
Чому фотосинтетична бактерія має різні пігменти?
Незалежні від світла реакції
Після того, як енергія сонця перетворюється в хімічну енергію і тимчасово зберігається в молекулах АТФ і НАДПГ (тривалість життя мільйонних часток секунди), фотоавтотрофи мають паливо, необхідне для побудови молекул багатовуглеводів, які можуть вижити протягом сотень мільйонів років, протягом тривалого часу накопичувач енергії. Вуглець надходить від CO 2, газу, який є відпрацьованим продуктом клітинного дихання.
Цикл Кальвіна-Бенсона (названий на честь Мелвіна Кальвіна [1911—1997] та Ендрю Бенсона [1917—2015]), біохімічний шлях, який використовується для фіксації CO 2, знаходиться в цитоплазмі фотосинтетичних бактерій та в стромі еукаріотичних хлоропластів. Світлонезалежні реакції циклу Кальвіна можуть бути організовані на три основні етапи: фіксація, зменшення та регенерація (докладну ілюстрацію циклу Кальвіна див. Додаток С).
- Фіксація: фермент рибулоза бісфосфаткарбоксилаза (RubiSCo) каталізує додавання СО 2 до бісфосфату рибулози (RubP). Це призводить до отримання 3-фосфогліцерату (3-PGA).
- Зменшення: Шість молекул як АТФ, так і NADPH (від світлозалежних реакцій) використовуються для перетворення 3-PGA в 3-фосфат гліцеральдегіду (G3P). Деякий G3P потім використовується для побудови глюкози.
- Регенерація: Решта G3P, що не використовується для синтезу глюкози, використовується для регенерації RubP, що дозволяє системі продовжувати фіксацію CO 2. Ще три молекули АТФ використовуються в цих реакціях регенерації.
Цикл Кальвіна широко використовується рослинами та фотоавтотрофними бактеріями, а фермент RubiSCo, як кажуть, є найбільш рясним ферментом на землі, що становить 30% - 50% загального розчинного білка в рослинних хлоропластах. 1 Однак, крім його поширеного використання у фотоавтотрофах, цикл Кальвіна також використовується багатьма нефотосинтетичними хемоавтотрофами для фіксації CO 2. Додатково інші бактерії та археї використовують альтернативні системи для фіксації СО 2. Хоча більшість бактерій, що використовують альтернативи циклу Кальвіна, є хемоавтотрофними, деякі зелені сірчані фотоавтотрофні бактерії також показали, що використовують альтернативний шлях фіксації CO 2.
Вправа\(\PageIndex{3}\)
Опишіть три етапи циклу Кальвіна.
Ключові поняття та резюме
- Гетеротрофи залежать від вуглеводів, що виробляються автотрофами, багато з яких є фотосинтетичними, перетворюючи сонячну енергію в хімічну енергію.
- Різні фотосинтезуючі організми використовують різні суміші фотосинтетичних пігментів, які збільшують діапазон довжин хвиль світла, який організм може поглинати.
- Фотосистеми (PSI і PSII) містять світлозбиральний комплекс, що складається з декількох білків і пов'язаних пігментів, які поглинають світлову енергію. Світлозалежні реакції фотосинтезу перетворюють сонячну енергію в хімічну енергію, виробляючи АТФ і NADPH або NADH для тимчасового зберігання цієї енергії.
- При кисневому фотосинтезі H 2 O служить донором електронів для заміни електрона реакційного центру, а кисень утворюється як побічний продукт. При аноксигенному фотосинтезі в якості донора електронів можуть використовуватися інші відновлені молекули, такі як H 2 S або тіосульфат; як такий, кисень не утворюється як побічний продукт.
- Нециклічне фотофосфорилювання використовується в кисневому фотосинтезі, коли є необхідність як у виробництві АТФ, так і NADPH. Якщо потреби клітини в АТФ перевищують її потреби в NADPH, то замість цього вона може проводити циклічне фотофосфорилювання, виробляючи лише АТФ.
- Світлонезалежні реакції фотосинтезу використовують АТФ і NADPH від світлозалежних реакцій для фіксації CO 2 в молекулах органічного цукру.
Виноски
- 1 А. Дхінгра та співавт. «Покращена трансляція гена Rbc S, експресованого хлоропластом, відновлює малі рівні субодиниці та фотосинтезу в ядерних рослинах-Rbc S Antisense.» Праці Національної академії наук Сполучених Штатів Америки 101 № 16 (2004) :6315—6320.