6.2: Життєвий цикл вірусу
- Page ID
- 3886

Цілі навчання
- Опишіть літичні та лізогенні життєві цикли
- Опишіть процес реплікації вірусів тварин
- Охарактеризуйте унікальні характеристики ретровірусів і латентних вірусів
- Обговоріть віруси людини та їх взаємодію між клітинами вірусу та господаря
- Поясніть процес трансдукції
- Охарактеризуйте процес реплікації вірусів рослин
Всі віруси залежать від клітин для розмноження і обмінних процесів. Самі по собі віруси не кодують всі ферменти, необхідні для реплікації вірусу. Але всередині клітини-господаря вірус може керувати клітинним апаратом, щоб виробляти більше вірусних частинок. Бактеріофаги розмножуються тільки в цитоплазмі, так як прокаріотичні клітини не мають ні ядра, ні органел. У еукаріотичних клітині більшість ДНК-вірусів можуть реплікуватися всередині ядра, за винятком спостерігаються у великих ДНК-вірусах, таких як поксвіруси, які можуть реплікуватися в цитоплазмі. РНК-віруси, які заражають клітини тварин, часто реплікуються в цитоплазмі.
Життєвий цикл вірусів з хостами прокаріотів
Життєвий цикл бактеріофагів був хорошою моделлю для розуміння того, як віруси впливають на клітини, які вони заражають, оскільки подібні процеси спостерігалися для вірусів еукаріотів, які можуть спричинити негайну загибель клітини або встановити приховану або хронічну інфекцію. Вірулентні фаги зазвичай призводять до загибелі клітини шляхом лізису клітин. З іншого боку, помірні фаги можуть стати частиною хромосоми господаря і реплікуються з геномом клітини до тих пір, поки їх не спонукають створювати знову зібрані віруси або віруси потомства.
Літичний цикл
Під час літичного циклу вірулентного фага бактеріофаг захоплює клітину, відтворює нові фаги, руйнує клітину. Т-навіть фаг є хорошим прикладом добре охарактеризованого класу вірулентних фагів. У літичному циклі бактеріофагів виділяють п'ять стадій (див. Рис.\(\PageIndex{1}\)). Приєднання - це перша стадія інфекційного процесу, при якій фаг взаємодіє зі специфічними рецепторами поверхні бактерій (наприклад, ліпополісахаридами та білком omPC на поверхнях господаря). Більшість фагів мають вузький діапазон господарів і можуть заражати один вид бактерій або один штам всередині виду. Це унікальне визнання може бути використано для цілеспрямованого лікування бактеріальної інфекції шляхом фагової терапії або для фагового типування для виявлення унікальних бактеріальних підвидів або штамів. Друга стадія зараження - проникнення або проникнення. Це відбувається через скорочення хвостової оболонки, яка діє як підшкірна голка для введення вірусного геному через клітинну стінку і мембрану. Головка фага і інші компоненти залишаються поза бактеріями.
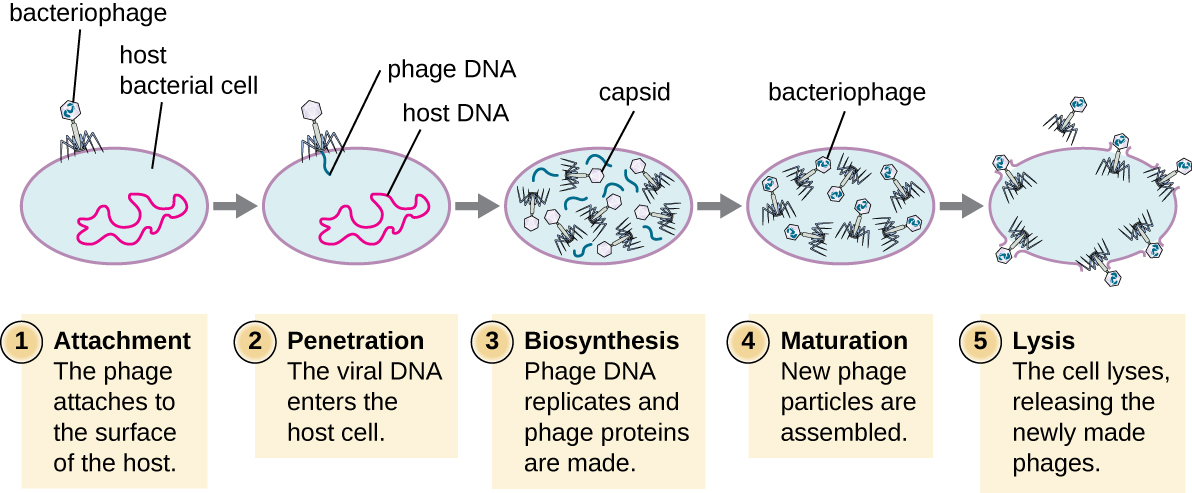
Третя стадія зараження - біосинтез нових вірусних компонентів. Після потрапляння в клітину-господар вірус синтезує закодовані вірусом ендонуклеази для деградації бактеріальної хромосоми. Потім він викрадає клітину господаря для реплікації, транскрибування та перекладу необхідних вірусних компонентів (капсомери, оболонка, базові пластини, хвостові волокна та вірусні ферменти) для складання нових вірусів. Гени полімерази зазвичай виражаються на початку циклу, тоді як капсидні і хвостові білки виражаються пізніше. Під час фази дозрівання створюються нові віріони. Для звільнення вільних фагів бактеріальна клітинна стінка порушується фаговими білками, такими як холін або лізоцим. Завершальний етап - реліз. Зрілі віруси вириваються з клітини-господаря в процесі, який називається лізисом, і віруси потомства звільняються в навколишнє середовище, щоб заразити нові клітини.
Лізогенний цикл
У лізогенному циклі геном фага також потрапляє в клітину через приєднання і проникнення. Яскравим прикладом фага з таким типом життєвого циклу є лямбда-фаг. Під час лізогенного циклу замість того, щоб вбивати господаря, геном фага інтегрується в бактеріальну хромосому і стає частиною господаря. Інтегрований геном фага називається профагом. Бактеріальний господар з профагом називається лізогеном. Процес, при якому бактерія заражається помірним фагом, називається лізогенезом. Для помірних фагів характерно бути латентним або неактивним всередині клітини. Оскільки бактерія відтворює свою хромосому, вона також відтворює ДНК фага і передає її новим дочірнім клітинам під час розмноження. Наявність фага може змінити фенотип бактерії, оскільки він може принести додаткові гени (наприклад, гени токсину, які можуть збільшити бактеріальну вірулентність). Ця зміна фенотипу господаря називається лізогенним перетворенням або фаговим перетворенням. Деякі бактерії, такі як Vibrio cholerae та Clostridium botulinum, менш вірулентні за відсутності профага. Фаги, що заражають ці бактерії, переносять гени токсину в своєму геному і посилюють вірулентність господаря, коли гени токсину експресуються. У разі V. cholera, закодований фагом токсин може викликати сильну діарею; у C. botulinum токсин може викликати параліч. Під час лізогенезу пророцтво зберігатиметься в хромосомі господаря до індукції, що призводить до висічення вірусного генома з хромосоми господаря. Після того, як відбулася індукція, помірний фаг може протікати через літичний цикл, а потім піддаватися лізогену у щойно інфікованій клітині (див. Рис.\(\PageIndex{2}\)).
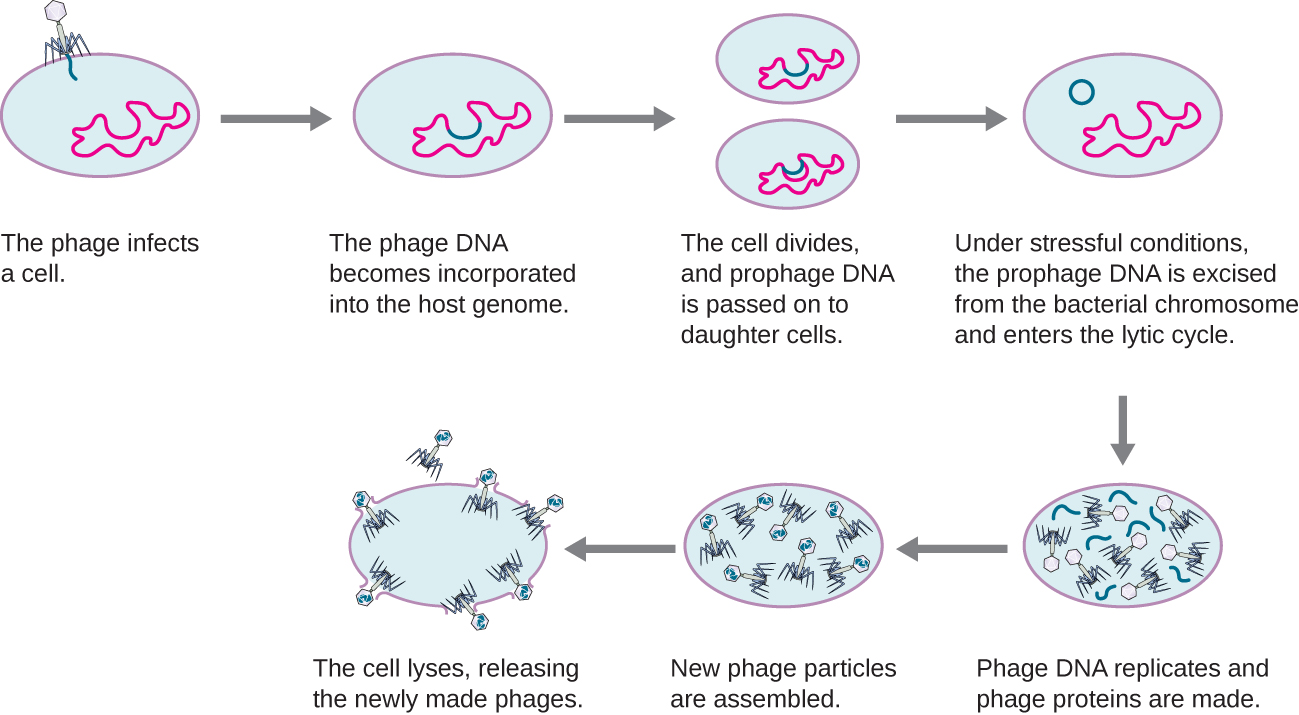
Це відео ілюструє етапи лізогенного життєвого циклу бактеріофага і перехід в літичну фазу.
Вправа\(\PageIndex{1}\)
Чи не можна виявити прихований фаг у бактерії?
Трансдукція
Трансдукція відбувається, коли бактеріофаг переносить бактеріальну ДНК від однієї бактерії до іншої під час послідовних інфекцій. Існує два види трансдукції: узагальнена і спеціалізована трансдукція. Під час літичного циклу вірусної реплікації вірус захоплює клітину-господаря, деградує хромосому господаря і робить більше вірусних геномів. Оскільки він збирає та упаковує ДНК у фагову головку, упаковка іноді робить помилку. Замість упаковки вірусної ДНК, він бере випадковий шматок ДНК господаря і вставляє його в капсид. Після звільнення цей віріон буде вводити ДНК колишнього господаря в нещодавно зараженого господаря. Безстатева передача генетичної інформації може дозволити відбутися рекомбінації ДНК, таким чином забезпечуючи нового господаря новими генами (наприклад, геном стійкості до антибіотиків або геном, що метаболізує цукор). Генералізована трансдукція відбувається, коли випадковий шматок бактеріальної хромосомної ДНК переноситься фагом під час літичного циклу. Спеціалізована трансдукція відбувається в кінці лізогенного циклу, коли пророцтво січуть і бактеріофаг вступає в літичний цикл. Оскільки фаг інтегрований у геном господаря, пророцтво може відтворюватися як частина господаря. Однак деякі умови (наприклад, вплив ультрафіолетового світла або хімічне опромінення) стимулюють профаг піддаватися індукції, змушуючи фаг висікати з генома, вступати в літичний цикл і виробляти нові фаги, щоб залишити клітини-господаря. Під час процесу висічення з хромосоми господаря фаг може зрідка видаляти деяку бактеріальну ДНК поблизу місця вірусної інтеграції. ДНК фага та господаря з одного кінця або обох кінців сайту інтеграції упаковуються в капсид і передаються новому, зараженому господареві. Оскільки ДНК, перенесена фагом, не випадково упаковується, а є специфічним шматочком ДНК поблизу місця інтеграції, цей механізм передачі генів називають спеціалізованою трансдукцією (див. Рис.\(\PageIndex{3}\)). Потім ДНК може рекомбінувати з хромосомою господаря, надаючи останній нові характеристики. Трансдукція, здається, відіграє важливу роль в еволюційному процесі бактерій, надаючи їм механізм безстатевого обміну генетичною інформацією.
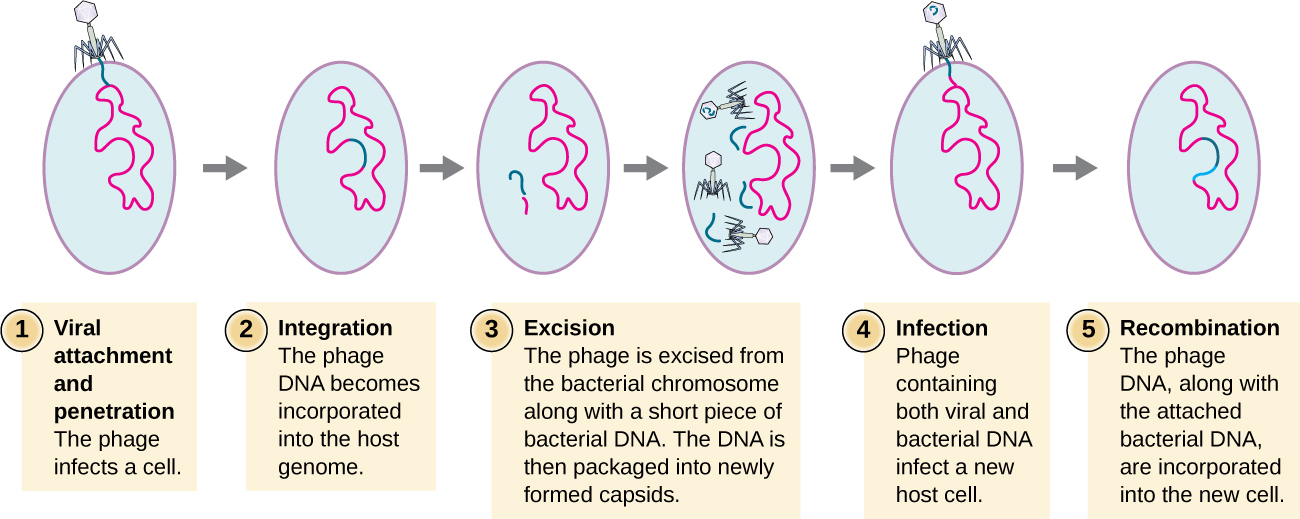
Вправа\(\PageIndex{2}\)
Який життєвий цикл фагів пов'язаний з якими формами трансдукції?
Життєвий цикл вірусів з тваринними господарями
Літичні віруси тварин слідують за аналогічними стадіями зараження бактеріофагами: приєднання, проникнення, біосинтез, дозрівання та вивільнення (див. Рис.\(\PageIndex{4}\)). Однак механізми проникнення, нуклеїновокислотного біосинтезу і вивільнення відрізняються між бактеріальними і тваринними вірусами. Після зв'язування з рецепторами-господарями віруси тварин потрапляють через ендоцитоз (поглинання клітиною-господарем) або через мембранний злиття (вірусна оболонка з мембраною клітини господаря). Багато вірусів є специфічними для господаря, тобто вони заражають лише певний тип господаря; і більшість вірусів заражають лише певні типи клітин всередині тканин. Така специфіка називається тканинним тропізмом. Приклади цього демонструє поліовірус, який проявляє тропізм для тканин головного і спинного мозку, або вірус грипу, який має первинний тропізм для дихальних шляхів.
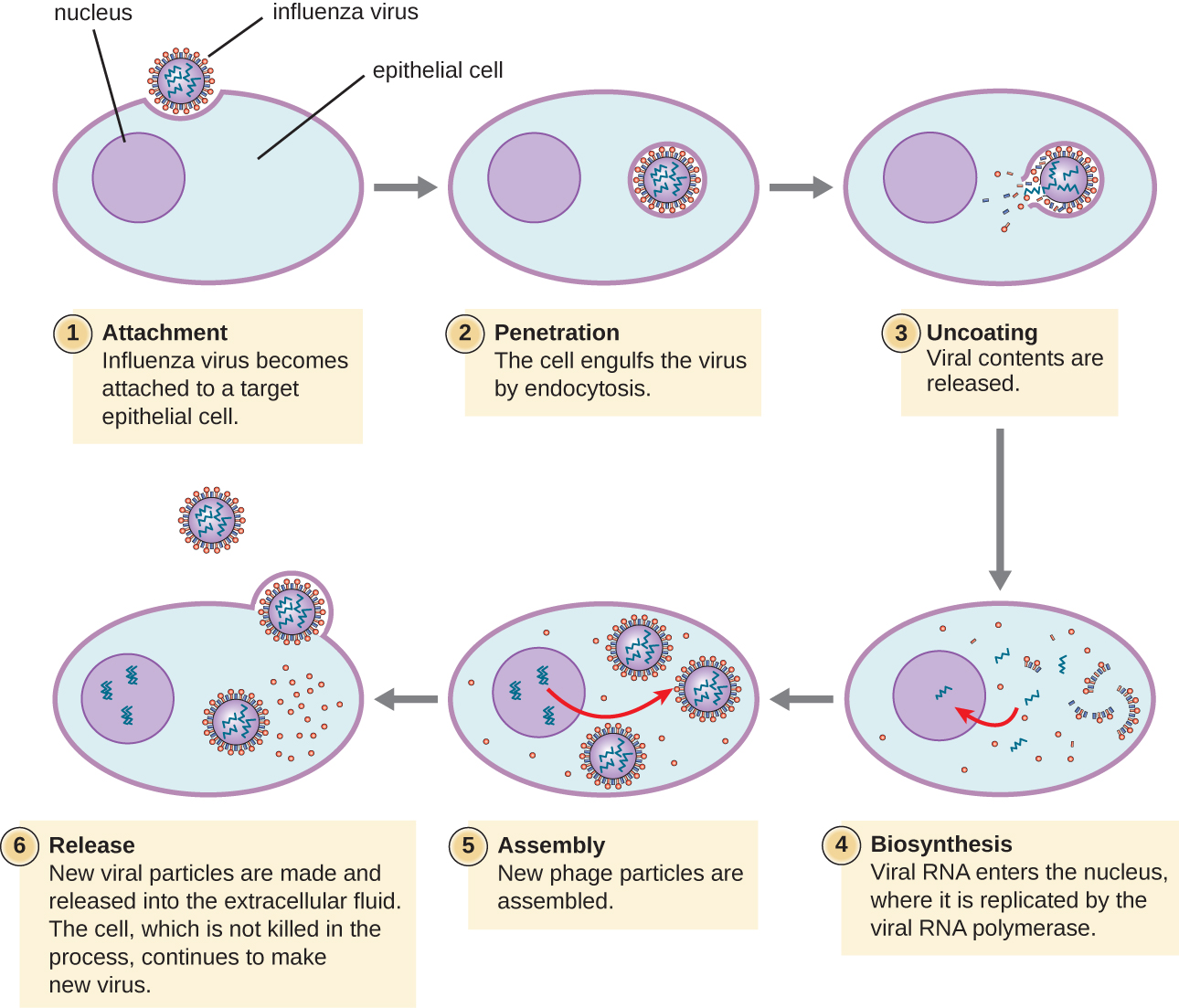
Віруси тварин не завжди експресують свої гени, використовуючи нормальний потік генетичної інформації - від ДНК до РНК до білка. Деякі віруси мають геном dsDNA, подібний до клітинних організмів, і можуть слідувати за нормальним потоком. Однак інші можуть мати геноми SsDNA, dSrNA або ssRNA. Характер генома визначає, як геном реплікується і виражається у вигляді вірусних білків. Якщо геном є SSDNA, ферменти господаря будуть використовуватися для синтезу другої нитки, яка доповнює нитку геному, таким чином виробляючи dsDNA. Тепер dsDNA можна реплікувати, транскрибувати та перекладати подібно до ДНК господаря.
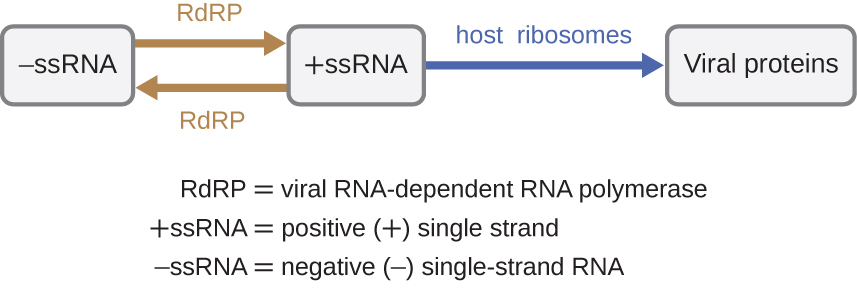
Якщо вірусним геном є РНК, необхідно використовувати інший механізм. Існує три типи генома РНК: dsRNA, позитивна (+) одножильна (+SsRNA) або негативна (−) одножильна РНК (−SSRNA). Якщо вірус має геном +SSRNA, його можна перевести безпосередньо, щоб зробити вірусні білки. Вірусна геномна+SSRNA діє як клітинна мРНК. Однак, якщо вірус містить геном −SSRNA, рибосоми господаря не можуть перевести його до тих пір, поки −SSRNA не буде репліковано на +SSRNA вірусною РНК-залежною РНК-полімеразою (rDRP) (див. Рисунок\(\PageIndex{5}\)). RdRP вводиться вірусом і може бути використано для створення +SSRNA з вихідного геному −SSRNA. RDRP також є важливим ферментом для реплікації вірусів dsRNA, оскільки він використовує негативну нитку двоцепочечного генома як шаблон для створення+SSRNA. Нещодавно синтезовані копії +SSRNA потім можуть бути переведені клітинними рибосомами.
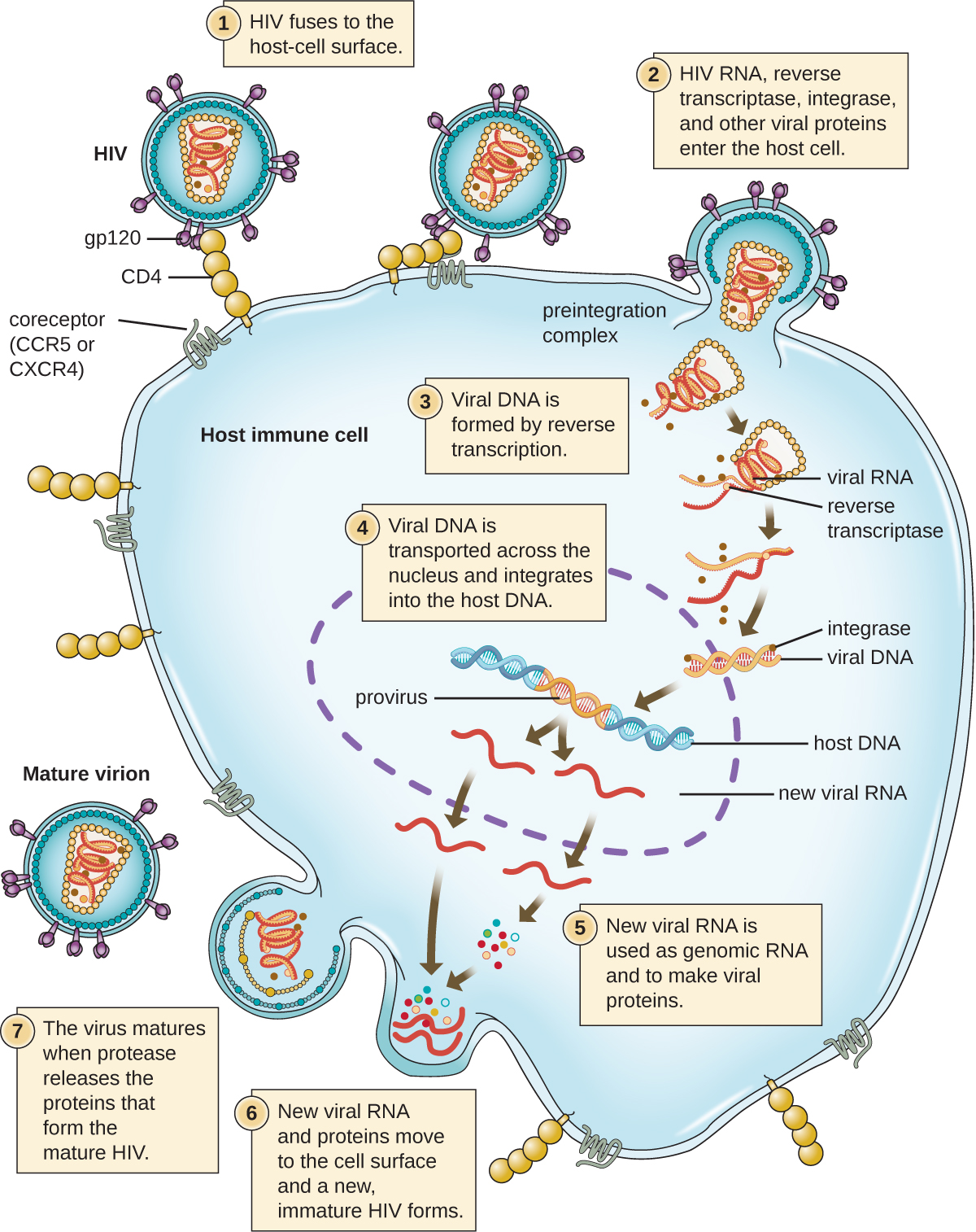
Альтернативний механізм синтезу вірусних нуклеїнових кислот спостерігається у ретровірусних ес, які представляють собою+віруси SSRNA (див. Рис.\(\PageIndex{6}\)). Одноланцюгові РНК-віруси, такі як ВІЛ, несуть спеціальний фермент, який називається зворотною транскриптазою всередині капсиду, який синтезує додаткову копію SSDNA (кДНК), використовуючи геном +ssRNA як шаблон. Потім SSDNA перетворюється в dsDNA, яка може інтегруватися в хромосому господаря і стати постійною частиною господаря. Інтегрований вірусний геном називається провірусом. Вірус тепер може залишатися в господареві протягом тривалого часу, щоб встановити хронічну інфекцію. Стадія провіруса схожа на стадію пророцтва при бактеріальній інфекції під час лізогенного циклу. Однак, на відміну від пророцтва, провирус не піддається висічення після зрощування в геном.
Вправа\(\PageIndex{3}\)
РНК-залежна РНК-полімераза виготовлена з вірусного гена або гена-господаря?
Персистуючі інфекції
Персистуюча інфекція виникає, коли вірус не повністю очищається від системи господаря, а залишається в певних тканині або органах інфікованої людини. Вірус може мовчати або піддаватися продуктивному зараженню, не завдаючи серйозної шкоди і не вбиваючи господаря. Механізми персистуючої інфекції можуть включати регуляцію вірусних або генних виразів господаря або зміну імунної відповіді господаря. Дві первинні категорії персистируючих інфекцій - латентна інфекція та хронічна інфекція. Приклади вірусів, що викликають латентні інфекції, включають вірус простого герпесу (оральний та генітальний герпес), вірус вітряної віспи (вітрянка та оперізуючий лишай) та вірус Епштейна-Барра (мононуклеоз). Вірус гепатиту С та ВІЛ - два приклади вірусів, які викликають тривалі хронічні інфекції.
латентна інфекція
Не всі віруси тварин піддаються реплікації літичним циклом. Існують віруси, які здатні залишатися прихованими або сплячими всередині клітини в процесі, який називається латентністю. Ці типи вірусів відомі як латентні віруси і можуть викликати приховані інфекції. Віруси, здатні до латентності, спочатку можуть викликати гостру інфекцію, перш ніж стати сплячим.
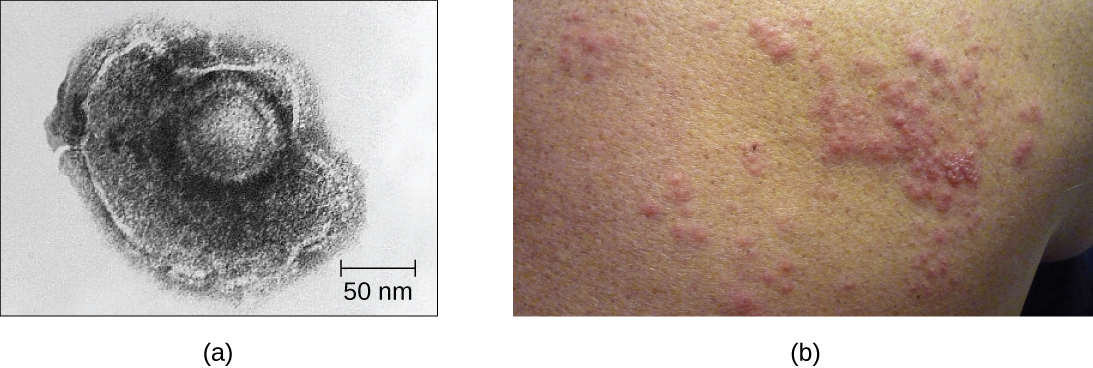
Наприклад, вірус вітряної віспи заражає багато клітин по всьому організму і викликає вітрянку, що характеризується висипом пухирів, що покривають шкіру. Приблизно від 10 до 12 днів постінфекції захворювання розсмоктується, і вірус переходить у сплячий стан, проживаючи в нервово-клітинних гангліях роками. За цей час вірус не вбиває нервові клітини і не продовжує розмножуватися. Незрозуміло, чому вірус перестає розмножуватися всередині нервових клітин і виражає мало вірусних білків, але в деяких випадках, як правило, після багатьох років спокою вірус реактивується і викликає нове захворювання під назвою оперізуючий лишай (рис.\(\PageIndex{7}\)). У той час як вітряна віспа вражає багато областей по всьому тілу, оперізуючий лишай - це специфічне захворювання нервових клітин, що виникає з гангліїв, в яких вірус був сплячим.
Латентні віруси можуть залишатися сплячими, існуючи як кругові молекули вірусного генома поза хромосомою господаря. Інші стають провірусами шляхом інтеграції в геном господаря. Під час спокою віруси не викликають жодних симптомів захворювання і можуть бути важко виявити. Пацієнт може не знати, що він або вона переносить вірус, якщо не було проведено вірусний діагностичний тест.
Хронічна інфекція
Хронічна інфекція - це захворювання з симптомами, які рецидивуючі або постійні протягом тривалого часу. Деякі вірусні інфекції можуть бути хронічними, якщо організм не в змозі усунути вірус. ВІЛ є прикладом вірусу, який виробляє хронічну інфекцію, часто після тривалого періоду латентності. Як тільки людина заражається ВІЛ, вірус може бути виявлений в тканині безперервно після цього, але неліковані пацієнти часто не відчувають симптомів протягом багатьох років. Однак вірус підтримує хронічну персистенцію за допомогою декількох механізмів, які перешкоджають імунній функції, включаючи запобігання експресії вірусних антигенів на поверхні інфікованих клітин, змінюючи самі імунні клітини, обмежуючи експресію вірусних генів та швидко змінюючи вірусні антигени через мутація. Зрештою, пошкодження імунної системи призводить до прогресування захворювання, що призводить до синдрому набутого імунодефіциту (СНІД). Різні механізми, які ВІЛ використовує, щоб уникнути очищення імунною системою, також використовуються іншими хронічно зараженими вірусами, включаючи вірус гепатиту С.
Вправа\(\PageIndex{4}\)
Якими двома способами вірусу вдається зберегти стійку інфекцію?
Життєвий цикл вірусів з рослинними хостами
Віруси рослин більше схожі на віруси тварин, ніж на бактеріофаги. Віруси рослин можуть бути обволікаються або не обволікаються. Як і багато вірусів тварин, рослинні віруси можуть мати або ДНК, або РНК геном і бути однонитковими або дволанцюжковими. Однак більшість рослинних вірусів не мають генома ДНК; більшість мають геном +SSRNA, який діє як месенджерна РНК (мРНК). Лише меншість рослинних вірусів мають інші типи геномів.
Віруси рослин можуть мати вузький або широкий діапазон господарів. Наприклад, вірус цитрусової трістези заражає лише кілька рослин роду Citrus, тоді як вірус огіркової мозаїки заражає тисячі рослин різних сімейств рослин. Більшість рослинних вірусів передається при контакті між рослинами, або грибами, нематодами, комахами або іншими членистоногими, які діють як механічні переносники. Однак деякі віруси можуть передаватися лише певним типом переносника комах; наприклад, певний вірус може передаватися попелиць, але не білокрилки. У деяких випадках віруси також можуть потрапляти в здорові рослини через рани, як це може статися через обрізку або пошкодження погоди.
Віруси, що вражають рослини, вважаються біотрофними паразитами, а значить, вони можуть встановити інфекцію, не вбиваючи господаря, подібно до того, що спостерігається в лізогенних життєвих циклах бактеріофагів. Вірусна інфекція може протікати безсимптомно (латентно) або може призвести до загибелі клітин (літична інфекція). Життєвий цикл починається з проникнення вірусу в клітину-господаря. Далі вірус не покривається всередині цитоплазми клітини при видаленні капсиду. Залежно від типу нуклеїнової кислоти клітинні компоненти використовуються для реплікації вірусного генома і синтезу вірусних білків для складання нових віріонів. Для встановлення системної інфекції вірус повинен потрапити в таку частину судинної системи рослини, як, наприклад, флоем. Час, необхідний для системної інфекції, може варіюватися від декількох днів до декількох тижнів залежно від вірусу, виду рослин та умов навколишнього середовища. Життєвий цикл вірусу завершений, коли він передається від зараженої рослини до здорової рослини.
Вправа\(\PageIndex{5}\)
Яка будова і геном типового рослинного вірусу?
Крива вірусного росту
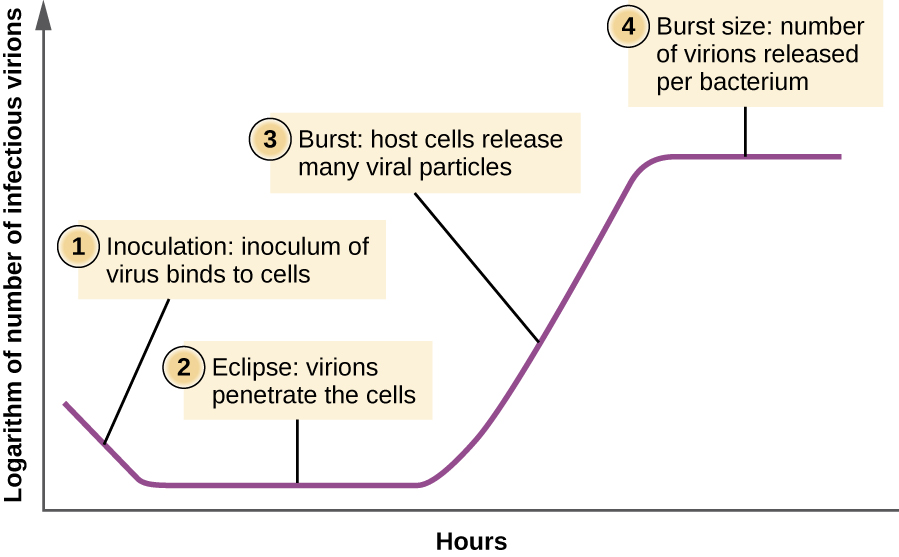
На відміну від кривої зростання бактеріальної популяції, крива зростання популяції вірусу протягом її життєвого циклу не слідує сигмоїдальній кривій. Під час початкової стадії щеплення вірусом викликає інфекцію. У фазі затемнення віруси зв'язуються і проникають в клітини без віріонів, виявлених в середовищі. Основна відмінність, яка наступна з'являється у кривій росту вірусу порівняно з кривою росту бактерій, виникає, коли віріони вивільняються з лізованої клітини-господаря одночасно. Таке явище називається вибухом, а кількість віріонів на одну бактерію, що виділяється, описується як розмір вибуху. У одноступінчастої кривій розмноження бактеріофага клітини-господаря лізуються, виділяючи багато вірусних частинок в середовище, що призводить до дуже крутого підйому вірусного титру (кількості віріонів на одиницю об'єму). Якщо життєздатних клітин господаря не залишається, вірусні частинки починають деградувати під час занепаду культури (див. Рис.\(\PageIndex{8}\)).
Вправа\(\PageIndex{6}\)
Який аспект життєвого циклу вірусу призводить до раптового збільшення кривої зростання?
Незареєстровані методи лікування
Ебола невиліковна і смертельна. Спалах у Західній Африці в 2014 році був безпрецедентним, карликуючи інші людські епідемії Еболи за рівнем смертності. З 24 666 підозрюваних або підтверджених випадків загинуло 10 179 осіб. 1
Немає затверджених методів лікування або вакцин проти Еболи. Хоча деякі препарати виявили потенціал у лабораторних дослідженнях та моделювання тварин, вони не були перевірені на людях на безпеку та ефективність. Ці препарати не тільки не перевірені або незареєстровані, але вони також не мають дефіциту.
Враховуючи великі страждання та високий рівень смертності, справедливо запитати, чи краще незареєстровані та неперевірені ліки, ніж взагалі відсутні. Чи слід відпускати такі препарати і, якщо так, то хто повинен їх отримувати, зважаючи на їх вкрай обмежені запаси? Чи етично лікувати неперевірені препарати у пацієнтів з Еболою? З іншого боку, чи етично утримувати потенційно рятувальні ліки від вмираючих пацієнтів? Або ліки, можливо, слід зарезервувати для медичних працівників, які працюють над стримуванням захворювання?
У серпні 2014 року двоє інфікованих працівників американської допомоги та іспанського священика лікувалися zMapp, незареєстрованим препаратом, який був протестований у мавп, але не на людях. Двоє американських працівників допомоги одужали, але священик помер. Пізніше того ж місяця ВООЗ опублікувала звіт про етику лікування пацієнтів препаратом. Оскільки Ебола часто смертельна, колегія аргументувала, що етично давати незареєстровані наркотики та неетично утримувати їх з міркувань безпеки. Ця ситуація є прикладом «співчутливого використання» поза усталеною системою регулювання та управління терапією.
Ебола в США
24 вересня 2014 року Томас Ерік Дункан прибув до техаської пресвітеріанської лікарні в Далласі зі скаргою на лихоманку, головний біль, блювоту та діарею - симптоми, які зазвичай спостерігаються у пацієнтів із застудою або грипом. Після огляду лікар відділення невідкладної допомоги поставив йому діагноз синусит, призначив деякі антибіотики і відправив додому. Через два дні Дункан повернувся в лікарню на швидкій допомозі. Його стан погіршився, і додаткові аналізи крові підтвердили, що він був заражений вірусом Ебола.
Подальші дослідження показали, що Дункан щойно повернувся з Ліберії, однієї з країн, що пережили важку епідемію Ебола. 15 вересня, за дев'ять днів до того, як він з'явився в лікарні в Далласі, Дункан допоміг транспортувати сусіда, ураженого Еболою, до лікарні в Ліберії. Лікарня продовжувала лікувати Дункана, але він помер через кілька днів після прийому.
Хронологія випадку Дункана свідчить про життєвий цикл вірусу Ебола. Час інкубації Ебола становить від 2 днів до 21 дня. Між впливом Дункана вірусної інфекції та появою його симптомів пройшло дев'ять днів. Це частково відповідає періоду затемнення в зростанні популяції вірусу. Під час фази затемнення Дункан не зміг би передати хворобу іншим. Однак, як тільки заражена людина починає проявляти симптоми, хвороба стає дуже заразною. Вірус Ебола передається при безпосередньому контакті з краплями тілесних рідин, таких як слина, кров та блювотні маси. Дункан міг передати хворобу іншим у будь-який час після того, як у нього почалися симптоми, імовірно, за деякий час до його прибуття до лікарні в Далласі. Після того, як лікарня усвідомлює, що такий пацієнт, як Дункан, заражений вірусом Ебола, пацієнта негайно поміщають на карантин, і чиновники охорони здоров'я ініціюють зворотний слід, щоб ідентифікувати всіх, з ким пацієнт, як Дункан, міг взаємодіяти протягом періоду, в якому він виявляв симптоми.
Чиновники охорони здоров'я змогли відстежити 10 осіб з високим ризиком (члени сім'ї Дункана) та 50 осіб з низьким рівнем ризику, щоб контролювати їх на наявність ознак інфекції. Ніхто не заразився хворобою. Однак одна з медсестер, звинувачених у догляді Дункана, заразилася. Це, поряд з початковим помилковим діагнозом Дункана, дало зрозуміти, що лікарні США повинні забезпечити додаткову підготовку медичного персоналу, щоб запобігти можливому спалаху Еболи в США.
Вправа\(\PageIndex{7}\)
- Які види навчання можуть підготувати медичних працівників до стримування нових епідемій, таких як спалах Ебола 2014 року?
- У чому різниця між заразним збудником і інфекційним збудником?

Для отримання додаткової інформації про Еболу, будь ласка, відвідайте веб-сайт CDC.
Резюме
- Багато вірусів націлені на конкретних господарів або тканин. Деякі можуть мати більше одного хоста.
- Багато вірусів проходять кілька етапів, щоб інфікувати клітини господаря. Ці етапи включають приєднання, проникнення, розкриття, біосинтез, дозрівання та вивільнення.
- Бактеріофаги мають літичний або лізогенний цикл. Літичний цикл призводить до загибелі господаря, тоді як лізогенний цикл призводить до інтеграції фага в геном господаря.
- Бактеріофаги вводять ДНК в клітину господаря, тоді як віруси тварин потрапляють шляхом ендоцитозу або злиття мембран.
- Віруси тварин можуть зазнавати латентність, подібну до лізогенезу для бактеріофага.
- Більшість рослинних вірусів є позитивно-нитковими SSRNA і можуть піддаватися затримці, хронічній або літичній інфекції, як це спостерігається для вірусів тварин.
- Крива росту популяцій бактеріофагів - це одноступінчаста крива розмноження, а не сигмоїдальна крива, порівняно з кривою росту бактерій.
- Бактеріофаги передають генетичну інформацію між господарями за допомогою генералізованої або спеціалізованої трансдукції.
Виноски
- 1 Всесвітня організація охорони здоров'я. «Дані та статистика Еболи ВООЗ». 18 березня 2005 року. http://apps.who.int/gho/data/view.eb...150318? lang=uk