2: Запліднення та обертання корки
- Page ID
- 3358

Запліднення
Для статево розмножуються видів завершення запліднення знаменує початок ембріогенезу. Ви вже знаєте, що види мають різні механізми репродуктивної ізоляції, які перешкоджають міжвидовому заплідненню. У випадку з морськими їжаками (посилання нижче) ви побачите, що існує кілька кроків, які можуть опосередковувати розпізнавання специфічних видів. До них відносяться хемоаттрактанти (хімічні речовини, що виділяються яйцеклітиною, на які реагує сперма) та липкі молекули, що утримуються спермою (біндіни), які взаємодіють лише з тими ж видами яйцеклітин.
Точка входу сперми також значна у багатьох видів тим, що вона знаменує перший розрив в симетрії яєць. Уявіть собі досить симетричний куля. Тепер намалюйте одну точку на цій кулі, ця точка тепер орієнтир. Ви можете бути навпроти точки, близько до точки, або прямо на точці. Точка входу сперми, як і точка, виступає орієнтиром для порушення симетрії. Він робить це принаймні двома спорідненими способами, пожертвуючи центросому яйцеклітині та викликаючи цитоскелетні перебудови. Центросома сперми, крихітна органела, яка діє як насіння для утворення мікротрубочок, може виступати організатором веретеночного апарату. Поставивши одну сторону веретена біля точки входу сперми, він може визначити вісь першого відколу (рис. 1).
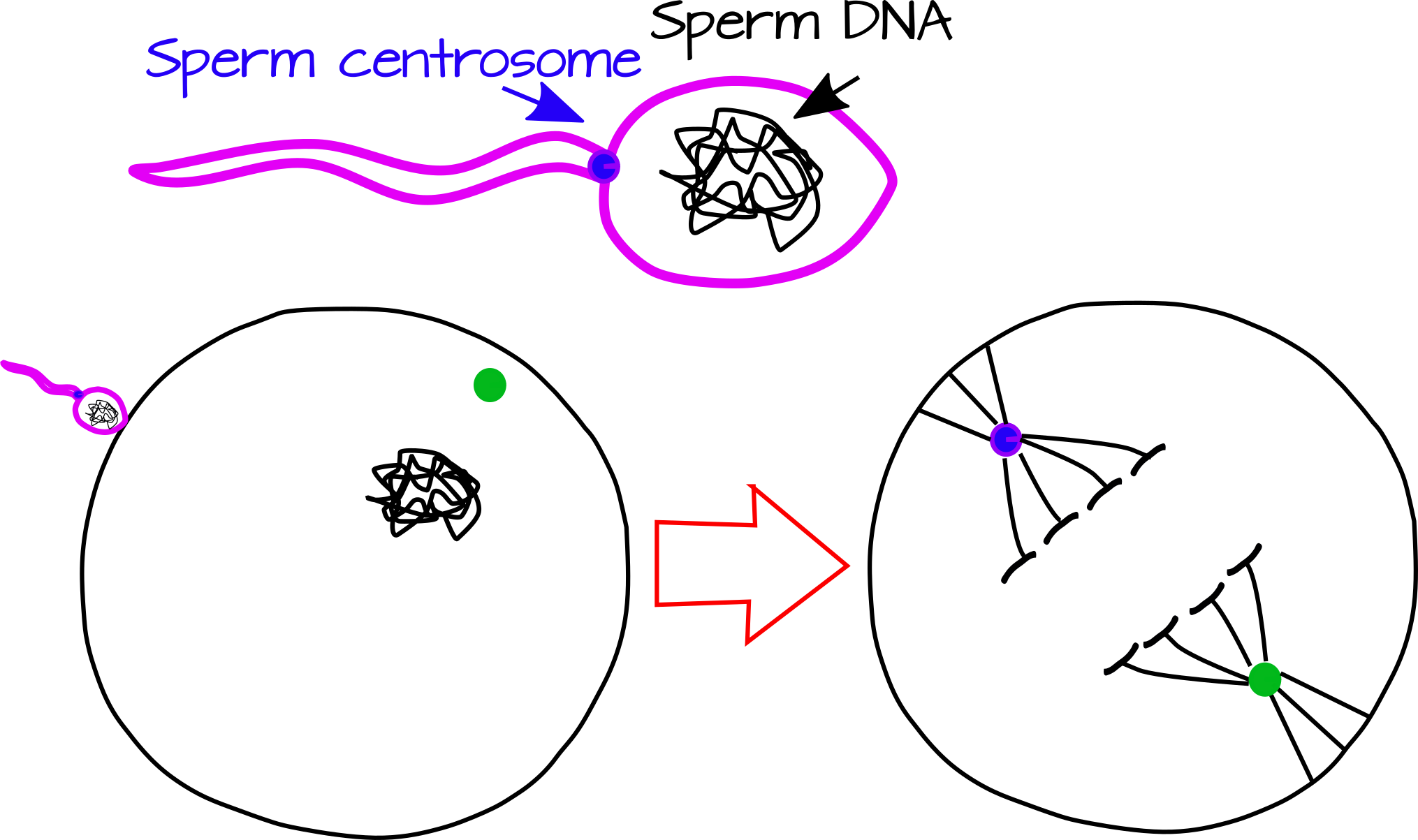
Крім того, точка входу сперми може бути тригером для цитоскелетного зсуву, включаючи процес, званий корковою ротацією, при якому зовнішній шар клітини (кора) обертається щодо внутрішньої цитоплазми. Хоча коркове обертання обмежене кількома видами (про які ми знаємо), вивчення його призвело до проривів у EvoDevo та біології розвитку загалом.
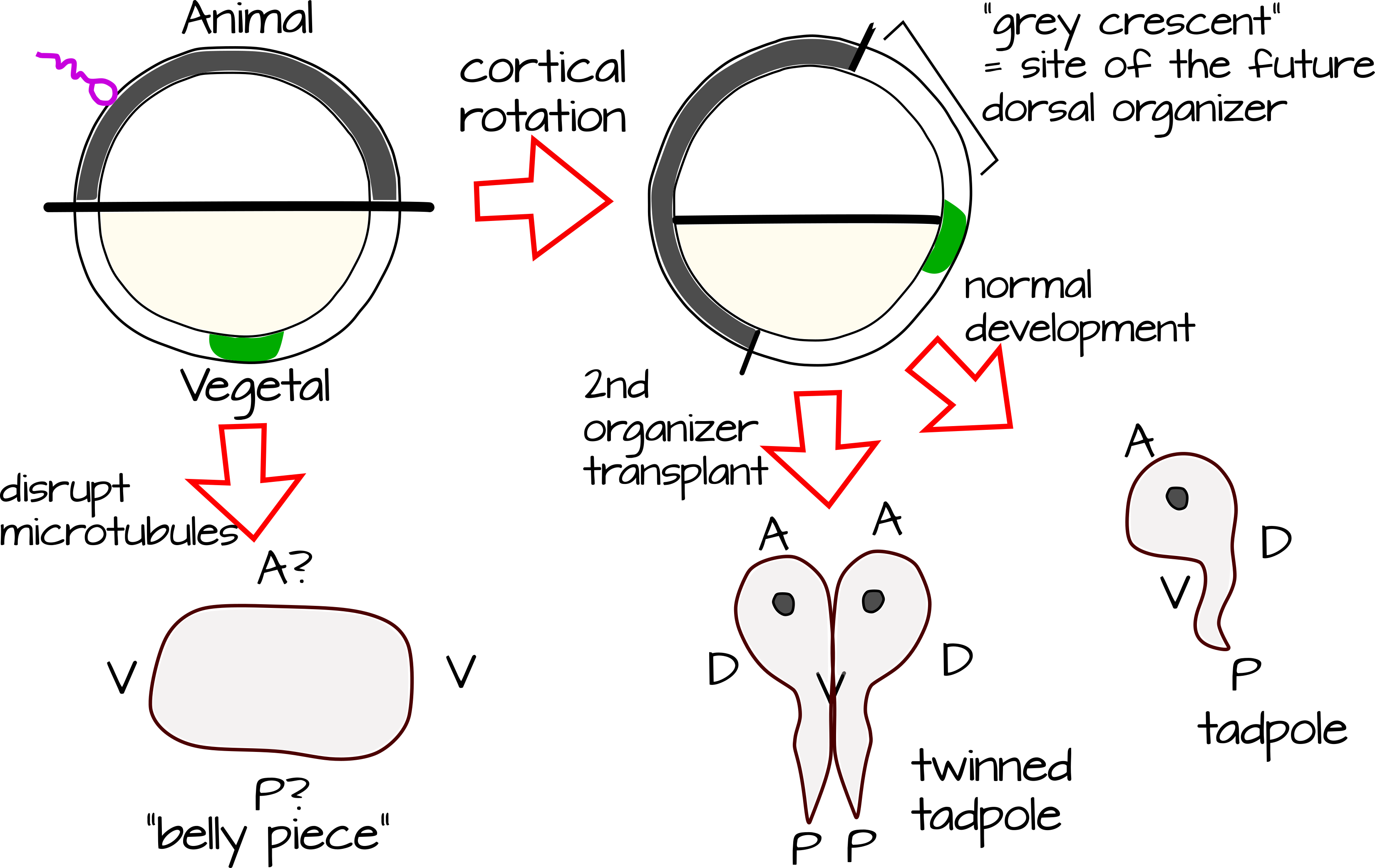
Значення коркового обертання як подія порушення симетрії підкреслюється, коли ми бачимо, що станеться, якщо ми запобігаємо цьому. Короткі методи лікування заплідненої яйцеклітини жаби (Xenopus laevis) агентами, які впливають на полімеризацію мікротрубочок, запобігають обертання корки і призводять до «вентралізованого» ембріона - тобто він має черевні структури, але не вистачає спинного структури, в тому числі ті, які б побудувати спинний мозок. Ми називаємо такий ембріон «шматочком живота». Що таке коркове обертання, яке перетворює жабу в жабу, а не в масу тканин живота?
Кортикальне обертання як приклад EvoDevo
Незважаючи на те, що коркове обертання - це процес, виявлений лише у кількох видів тварин, це приклад кількох фундаментальних концепцій у EvoDevo
1) Створення організаторів. Організатори - це частини ембріона, які виробляють сигнали, що малюють сусідні або віддалені тканини. Наприклад, у дрозофіл бікоїдна мРНК позначає органайзер голови. Виснаження бікоїдів призводить до безголових мух, занадто велика кількість бікоїдів призводить до великоголових личинок. Пізніше в семестрі ми розглянемо організатор ZPA в кінцівках хребетних.
Жаби мають організатора Spemann-Mangold (названий на честь студентки Хільди Мангольд та її радника Ганса Спемана, який відкрив та охарактеризував його) 2. Видалення тканини органайзера призводить до утворення кульки тканини живота. Додавання органайзера в інше місце на ембріоні призводить до побратимів пуголовків.
2) Індукція клітинної диференціації. Багато комірок мають шлях «за замовчуванням», який вони будуть слідувати з часом, якщо їм не будуть надані додаткові вказівки. Однією з головних ролей організатора є створення та надсилання цих додаткових інструкцій. Організатори роблять це за допомогою клітинних сигнальних молекул та регуляції генів. Приклад бікоїдів у дрозофіли є прикладом регуляції генів. Приклад жаби демонструє як регуляцію генів, так і клітинну сигналізацію, як я незабаром опишу.
3) Розвиток відбувається в 4D. Ми не тільки повинні думати про складні 3D-об'єкти, коли ми думаємо про розвиток, але ми також повинні враховувати час. З плином часу клітини і частини клітин змінюють свої положення. Це змінює «місцеве середовище» - зміна типів взаємодій, які можуть відбуватися. Це буде важливіше в гаструляції, але ми побачимо початок його тут у кортикальному обертанні. У разі коркового обертання обертання змінює місця розташування мРНК і білків. Вони в кінцевому підсумку будуть успадковані різними клітинами і дадуть цим клітинам початкові інструкції для диференціації. Ці клітини згодом змінять своє положення і в міру контакту з новими сусідами будуть далі диференціюватися і видавати нові вказівки.
Входження сперми викликає зміни в яйцеклітині
Незапліднена жаб'яча яйцеклітина не ідеально симетрична. Він має радіальну симетрію, тобто ви можете приклеїти шпильку зверху вниз і вона буде симетричною навколо шпильки. Однак верхня і нижня сторони відрізняються. Дно (рослинний полюс) збагачене щільними гранулами жовтка, а верх (полюс тварини) збагачений менш щільною цитоплазмою і пігментом. Подібна тварина/рослинна симетрія спостерігається у багатьох видів. Яйця жаби також асиметричні тим, що зроблені з двох шарів. Зовнішня кора і внутрішня клітинна маса. Тваринний та рослинний полюси відрізняються за мРНК та білками (див. Коротко, Генетика для оновлення термінами молекулярної біології) 3, а також внутрішня та зовнішня частини яйцеклітини.
Входження сперми викликає зміну способу взаємодії цитоскелетних мікротрубочок між собою. Вони переходять від неорганізованої мережі через клітину до паралельного масиву з найвищою щільністю в корі та на кордоні маси корки/внутрішньої клітини. Ці мікротрубочки обертають кору щодо внутрішньої клітинної маси, захоплюючи з собою асиметрично локалізовані коркові детермінанти 2. Оскільки пігментація в яйці була розподілена нерівномірно, це створює «сірий півмісяць», де рослинна кора тепер лежить над внутрішньою клітинною масою тварини (рис. 2)
Взаємодія між молекулярними детермінантами вегетальної кори та молекулярними детермінантами маси внутрішніх клітин тварин робить цей сірий півмісяць унікальним порівняно з тваринним або рослинним полюсом заплідненої яйцеклітини. Сірий півмісяць позначає майбутню спинну сторону жаби. Тепер ви можете зрозуміти, чому усунення коркового обертання або усунення молекулярних детермінант, що населяють сірий півмісяць, призводять до вентралізації жаби. Дублювання сірого півмісяця шляхом трансплантації або генетичної маніпуляції призводить до додаткової спинної осі і «побратимства» жаби.
У наступному розділі «Розщеплення та гаструляція» ми розглянемо розвиток та генетику цього організатора і подумаємо про наслідки організації тканин в еволюції.
Посилання
- «Мітоз у ембріона людини: життєво важлива роль центросоми сперми (центриоли)», 1997, Гістологія та гістопатологія, Генрі Сатанантан PMID:
9225167 - «Індукція ембріональної Примордії шляхом імплантації організаторів з різних видів», 1924, Архів мікроскопічної анатомії und Entwicklungsmechanik, Ганс Спеманн та Гільда Мангольд., PMID: 11291841
- Переглянуто в: «Кортикальне обертання та локалізація РНК месенджера при утворенні осі Xenopus», 2012, Біологія розвитку Wires, Дуглас Х'юстон doi: 10.1002/wdev.29
Подальше читання: Біологія розвитку
www.ncbi.nlm.nih.gov/Книги/NBK9983/
- Введення в запліднення
- 1. сперма і яйцеклітина
- 2. Як ви робите? Розпізнавання сперми та яєць
- 3. Один сперматозоїд на яйцеклітину: реакція коркової гранули
- 4. НЕОБОВ'ЯЗКОВО: Активація метаболізму яєць
- 5. НЕОБОВ'ЯЗКОВО: Два стають одним, злиття генетичного матеріалу
- 6. Порушення симетрії: перебудова цитоплазми яйцеклітини
- 7. Резюме