16.2: Еукаріотична епігенетична регуляція
- Page ID
- 8197
Модифікації ДНК, які не змінюють послідовність ДНК, можуть впливати на активність генів. Хімічні сполуки, які додаються до окремих генів, можуть регулювати їх активність; ці модифікації відомі як епігенетичні зміни. Епігеном включає всі хімічні сполуки, які були додані до всієї ДНК (геному) як спосіб регулювати активність (експресію) всіх генів у геномі. Хімічні сполуки епігенома не є частиною послідовності ДНК, а знаходяться на ДНК або прикріплені до неї («epi-» означає вище в грецькій мові). Епігеномні модифікації залишаються в міру поділу клітин і в деяких випадках можуть бути успадковані через покоління. Вплив навколишнього середовища, такі як дієта людини та вплив забруднюючих речовин, також можуть впливати на епігеном.
Епігенетичні зміни можуть допомогти визначити, чи включаються чи вимкнені гени, і можуть впливати на вироблення білків у певних клітині, гарантуючи, що виробляються лише необхідні білки. Наприклад, білки, що сприяють зростанню кісток, не виробляються в м'язових клітині. Шаблони модифікації епігеному різняться між окремими особами, різними тканинами всередині людини і навіть різними клітинами.
Регулювання доступу до генів
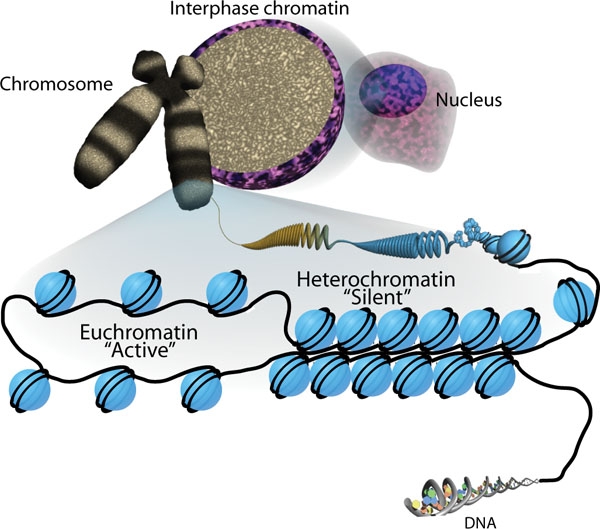
Геном людини кодує понад 20 000 генів, а це означає, що кожна з 23 пар хромосом людини містить тисячі генів. ДНК в ядрі кожної клітини точно намотана, згорнута і ущільнена в хромосоми так, що вона поміститься всередині ядерної мембрани. Він також організований таким чином, що конкретні сегменти можуть бути доступні в міру необхідності для певного типу комірки.
Перший рівень організації, або упаковки, - це обмотка ниток ДНК навколо гістонових білків. Гістони упаковують і впорядковують ДНК в структурні одиниці, звані нуклеосомними комплексами, які можуть контролювати доступ білків до областей ДНК (рис\(\PageIndex{1}\). Під електронним мікроскопом ця обмотка ДНК навколо гістонових білків з утворенням нуклеосом виглядає як маленькі кульки на нитці (рис.\(\PageIndex{1}\) Б). Ці кульки (білки гістону) можуть переміщатися по нитці (ДНК) і змінювати структуру молекули.

Якщо ДНК, що кодує конкретний ген, транскрибується в РНК, нуклеосоми, що оточують цю область ДНК, можуть ковзати вниз по ДНК, щоб відкрити цю специфічну хромосомну область і дозволити транскрипційним механізмам (РНК-полімеразі) ініціювати транскрипцію. Нуклеосоми можуть рухатися, щоб відкрити структуру хромосоми, щоб оголити сегмент ДНК, але робити це дуже контрольовано.
Активні відкриті ділянки хроматину називаються еухроматином (рис.\(\PageIndex{2}\)). Регіони генома, які транскрипційно активні, зазвичай евхроматичні. Щільно намотані ділянки хроматину називаються гетерохроматином. Гетерохроматичні області генома, як правило, замовчуються і транскрипційно неактивні.
Модифікації ДНК та гістонів
Як рухаються білки гістонів, і чи ДНК вільно або щільно обгорнута навколо них, залежить від сигналів, знайдених як на білках гістонів, так і на ДНК. Ці сигнали - це хімічні мітки, додані до білків гістонів та ДНК, які повідомляють гістонам, якщо хромосомна область повинна бути відкритою або закритою. Ці теги не є постійними, але їх можна додавати або видаляти за потребою. Вони являють собою хімічні модифікації (фосфатні, метилові або ацетильні групи), які приєднуються до специфічних амінокислот в білку або до нуклеотидів ДНК. Мітки не змінюють базову послідовність ДНК, але вони змінюють, наскільки щільно намотана ДНК навколо білків гістонів.
Цей тип генної регуляції називається епігенетичною регуляцією. Епігенетичний означає «навколо або вище генетики». Зміни, що відбуваються з білками гістону та ДНК, не змінюють послідовність нуклеотидів і не є постійними. Натомість ці зміни є тимчасовими, хоча вони можуть і часто зберігаються через кілька раундів поділу клітин. Вони змінюють хромосомну структуру (відкритий еухроматин або закритий гетерохроматин) у міру необхідності, але не змінюють послідовність основ всередині ДНК.
Ген може бути включений або вимкнений залежно від розташування та модифікацій білків гістонів та ДНК. Якщо ген повинен бути транскрибований, білки гістонів і ДНК модифікуються навколо хромосомної області, що кодує цей ген. Це відкриває хромосомну область (вона стає евхроматичною), щоб забезпечити доступ до РНК-полімерази та інших білків, званих факторами транскрипції, для зв'язування з промоторною областю, розташованою безпосередньо вище за течією гена, і ініціювати транскрипцію. Якщо ген залишиться вимкненим або замовченим, білки гістону та ДНК мають різні модифікації, які сигналізують про закриту хромосомну конфігурацію. У цій замкнутій конфігурації (гетерохроматин) РНК-полімераза та фактори транскрипції не мають доступу до ДНК і транскрипція не може відбуватися (рис.\(\PageIndex{2}\)).
Метилювання ДНК
Поширений тип епігеномної модифікації називається метилювання. Метилювання передбачає приєднання невеликих молекул, званих метильними групами, кожна з яких складається з одного атома вуглецю та трьох атомів водню, до нуклеотидів ДНК або амінокислот, що входять до складу білки гістону.
Коли ДНК метильована, метильна група зазвичай додається до нуклеотидів цитозину. Це відбувається в дуже специфічних регіонах, які називаються островами CpG. Це розтяжки з високою частотою цитозину і гуанін-динуклеотидних пар ДНК (CG), виявлених в промоторних областях генів. Коли така конфігурація існує, цитозиновий член пари може бути метильований (додається метильна група). Ця модифікація змінює спосіб взаємодії ДНК з білками, включаючи білки гістонів, які контролюють доступ до регіону. Коли метильні групи додаються до певного гена, цей ген вимикається або замовчується, і з цього гена не виробляється білок (рис.\(\PageIndex{3}\)).
Гіпотеза «Гістонового коду»
Гіпотеза гістонового коду - це гіпотеза про те, що транскрипція гена частково регулюється модифікаціями, внесеними в білки гістонів, насамперед на їх дещо гнучких кінцях (їх «хвостах»). Багато модифікацій хвоста гістону дуже добре корелюють зі структурою хроматину, і як стан модифікації гістону, так і структура хроматину добре корелюють з рівнями експресії генів. Найважливішою концепцією гіпотези гістонового коду є те, що модифікації гістонів служать для набору інших білків шляхом специфічного розпізнавання модифікованого гістону, а не просто стабілізуючи або дестабілізуючи взаємодію між гістоном та основною ДНК. Потім ці набрані білки діють, щоб активно змінювати структуру хроматину або сприяти транскрипції.
Код гістонів має потенціал бути масово складним. Є щонайменше 20 модифікацій, які зроблені до гістональних хвостів, які були відносно добре охарактеризовані, і є потенціал для багатьох інших, які ми не виявили. Кожен гістон може бути модифікований на декількох амінокислотах, з декількома різними хімічними модифікаціями. Інформація, яка може зберігатися в коді гістону, затьмарює кількість, яка зберігається в порядку основ в геномі людини.
Метлірування гістону
Частина білка гістону, відомого як хвіст гістону, може мати до нього метильні групи (CH 3). Це та сама модифікація, яка вноситься до нуклеотидів цитозину в ДНК. Специфічна амінокислота в хвості гістону, яка отримує метильований, дуже важлива для визначення того, чи буде вона затягувати або послабити структуру хроматину. Модифікація декількох амінокислот в хвості корелює з евхроматином і активною транскрипцією, в той час як модифікація на інші амінокислоти корелює з гетерохроматином і глушінням генів. Ви повинні знати, що гістони можуть бути метильованими, але ми не можемо використовувати метилювання гістону як предиктор для еухроматину або гетерохроматину.
Гістон ацетилювання
Гістонові хвости також можуть бути модифіковані додаванням ацетильної групи (цей процес відомий як ацетилювання гістону). Якщо ви пам'ятаєте з клітинного дихання, ацетильна група (така, що міститься в ацетил-КоА) - це 2-вуглецева молекула. Коли хвости гістону ацетильовані, це, як правило, призводить до розпушування хвостів навколо ДНК, дозволяючи хроматину розпушити (рис.\(\PageIndex{3}\)).

Інші модифікації
Існує багато інших модифікацій, які можуть бути внесені до гістонових білків на додаток до метилювання та ацетилювання. Хвости гістону можуть бути фосфорильованими або всюдисущими (де прикріплений невеликий білок під назвою убиквітин). Фосфорилювання гістону, здається, пов'язане з відновленням ДНК. Показано, що убіквітинація пов'язана як з активацією транскрипції, так і з інактивацією, залежно від конкретного місця.
Епігенетичні зміни
Оскільки помилки в епігенетичному процесі, такі як модифікація неправильного гена або неможливість додавання сполуки до гена, можуть призвести до аномальної активності генів або бездіяльності, вони можуть спричинити генетичні порушення. Виявлено, що стани, включаючи рак, порушення обміну речовин та дегенеративні розлади, пов'язані з епігенетичними помилками.
Ракові клітини часто мають ділянки ДНК, які показують різний рівень метилювання порівняно з нормальними клітинами. Деякі гени метилюються і замовчуються в ракових клітині, в той час як вони неметильовані і активні в нормальних клітині. Інші гени активні в ракових клітині, але неактивні в нормальних клітині. Кожен конкретний рак у кожної конкретної людини може демонструвати різні закономірності метилювання, хоча існують подібності між багатьма різними типами раку.
Вчені продовжують досліджувати взаємозв'язок між геномом і хімічними сполуками, які його модифікують. Зокрема, вони вивчають вплив модифікацій на функцію генів, вироблення білка та здоров'я людини.

Посилання
Якщо не зазначено інше, зображення на цій сторінці ліцензуються відповідно до CC-BY 4.0 OpenStax.
OpenStax, Концепції біології. OpenStax CNX. Травень 18, 2016 http://cnx.org/contents/b3c1e1d2-839...9a8aafbdd@9.10