4.1: Енергія та метаболізм


- Last updated
- Aug 3, 2022
- Save as PDF

Вчені використовують термін біоенергетика для опису поняття потоку енергії (рис 4.1.1) через живі системи, такі як клітини. Клітинні процеси, такі як побудова та руйнування складних молекул, відбуваються за допомогою ступінчастих хімічних реакцій. Деякі з цих хімічних реакцій є спонтанними і вивільняють енергію, тоді як інші вимагають енергії для продовження. Подібно до того, як живі істоти повинні постійно споживати їжу, щоб поповнити свої запаси енергії, клітини повинні постійно виробляти більше енергії для поповнення, що використовується багатьма потребують енергії хімічних реакцій, які постійно відбуваються. Разом всі хімічні реакції, що відбуваються всередині клітин, включаючи ті, які споживають або виробляють енергію, називаються метаболізмом клітини.
Метаболічні шляхи
Розглянемо метаболізм цукру. Це класичний приклад одного з численних клітинних процесів, які використовують і виробляють енергію. Живі істоти споживають цукру як основне джерело енергії, оскільки молекули цукру мають велику кількість енергії, що зберігається в їх зв'язках. Здебільшого фотосинтезуючі організми, такі як рослини, виробляють ці цукру. Під час фотосинтезу рослини використовують енергію (спочатку з сонячного світла) для перетворення вуглекислого газу (СО2) в молекули цукру (як глюкоза: C6H12O6). Вони споживають вуглекислий газ і виробляють кисень як продукт життєдіяльності. Ця реакція підсумовується як:
6CO2+6H2O⟶C6H12O6+6O2
Оскільки цей процес включає синтез молекули, що зберігає енергію, він вимагає введення енергії, щоб продовжити. Під час світлових реакцій фотосинтезу енергія забезпечується молекулою під назвою аденозинтрифосфат (АТФ), яка є первинною енергетичною валютою всіх клітин. Подібно до того, як долар використовується як валюта для придбання товарів, клітини використовують молекули АТФ як енергетичну валюту для виконання негайної роботи. На відміну від цього, молекули накопичення енергії, такі як глюкоза, витрачаються лише для того, щоб їх розщеплювати, щоб використовувати свою енергію. Реакцію, яка збирає енергію молекули цукру в клітинам, які потребують кисню для виживання, можна підсумувати зворотною реакцією на фотосинтез. У цій реакції витрачається кисень і виділяється вуглекислий газ як продукт життєдіяльності. Реакція підсумовується як:
C6H12O6+6O2⟶6H2O+6CO2
Обидві ці реакції передбачають багато кроків.
Процеси виготовлення та розщеплення молекул цукру ілюструють два приклади метаболічних шляхів. Метаболічний шлях - це низка хімічних реакцій, яка приймає вихідну молекулу і поступово модифікує її через низку проміжних метаболічних продуктів, врешті-решт отримуючи кінцевий продукт. На прикладі метаболізму цукру перший метаболічний шлях синтезував цукор з менших молекул, а інший шлях розбив цукор на більш дрібні молекули. Ці два протилежні процеси - перший, що вимагає енергії, а другий виробляє енергію - називаються анаболічними шляхами (будівельні полімери) та катаболічними шляхами (розщеплення полімерів на їх мономери) відповідно. Отже, метаболізм складається з синтезу (анаболізму) і деградації (катаболізму) (рис. 4.1.2).
Важливо знати, що хімічні реакції метаболічних шляхів не проходять самі по собі. Кожен етап реакції полегшується або каталізується білком, званим ферментом. Ферменти важливі для каталізації всіх типів біологічних реакцій - тих, які потребують енергії, а також тих, що виділяють енергію.
Енергія
Термодинаміка відноситься до вивчення передачі енергії та енергії за участю фізичної речовини. Матерія, що стосується конкретного випадку передачі енергії, називається системою, і все, що знаходиться поза цією матерією, називається оточенням. Наприклад, при нагріванні каструлі з водою на плиті система включає плиту, каструлю та воду. Енергія передається всередині системи (між плитою, каструлею і водою). Існує два типи систем: відкрита і закрита. У відкритій системі енергією можна обмінюватися з навколишнім середовищем. Система плити відкрита, оскільки тепло може втрачатися повітрю. Закрита система не може обмінюватися енергією з навколишнім середовищем.
Біологічні організми - це відкриті системи. Енергія обмінюється між ними та їх оточенням, оскільки вони використовують енергію сонця для фотосинтезу або споживають молекули, що зберігають енергію, і вивільняють енергію в навколишнє середовище, виконуючи роботу та виділяючи тепло. Як і всі речі у фізичному світі, енергія підпорядковується фізичним законам. Закони термодинаміки регулюють передачу енергії у всіх системах Всесвіту і між ними.
Взагалі, енергія визначається як здатність виконувати роботу, або створювати якісь зміни. Енергія існує в різних формах. Наприклад, електрична енергія, світлова енергія та теплова енергія - це різні типи енергії. Щоб оцінити те, як енергія надходить у біологічні системи та з них, важливо зрозуміти два фізичні закони, які керують енергією.
Термодинаміка
Перший закон термодинаміки говорить, що загальна кількість енергії у Всесвіті є постійним і законсервованим. Іншими словами, завжди було, і завжди буде, точно така ж кількість енергії у Всесвіті. Енергія існує в самих різних формах. Згідно з першим законом термодинаміки, енергія може передаватися з місця на місце або трансформуватися в різні форми, але вона не може бути створена або знищена. Передачі і перетворення енергії відбуваються навколо нас весь час. Лампочки перетворюють електричну енергію в світлову і теплову. Газові плити перетворюють хімічну енергію з природного газу в теплову. Рослини виконують одне з найбільш біологічно корисних перетворень енергії на землі: перетворення енергії сонячного світла в хімічну енергію, що зберігається в органічних молекул (рис 4.1.1). Деякі приклади енергетичних перетворень наведені на рисунку 4.1.3.
Завдання для всіх живих організмів полягає в тому, щоб отримати енергію з їх оточення у формах, які вони можуть передавати або перетворювати в корисну енергію для виконання роботи. Живі клітини еволюціонували, щоб вирішити цю проблему. Хімічна енергія, що зберігається в органічних молекулах, таких як цукру і жири, передається і перетворюється через ряд клітинних хімічних реакцій в енергію всередині молекул АТФ. Енергія в молекулах АТФ легко доступна для виконання роботи. Приклади видів робіт, які клітинам потрібно зробити, включають будівництво складних молекул, транспортування матеріалів, живлення руху вій або джгутиків та скорочення м'язових волокон для створення руху.
Основні завдання живої клітини з отримання, перетворення та використання енергії для виконання роботи можуть здатися простими. Однак другий закон термодинаміки пояснює, чому ці завдання складніше, ніж вони з'являються. Всі передачі енергії і перетворення ніколи не є повністю ефективними. У кожній передачі енергії деяка кількість енергії втрачається в непридатній для використання формі. У більшості випадків такою формою є теплова енергія. Термодинамічно теплова енергія визначається як енергія, що передається від однієї системи до іншої, яка не є робочою. Наприклад, при включенні лампочки частина енергії, що перетворюється з електричної в світлову, втрачається у вигляді теплової енергії. Так само частина енергії втрачається як теплова енергія під час клітинних метаболічних реакцій.
Важливим поняттям у фізичних системах є порядок і розлад. Чим більше енергії, яка втрачається системою в її оточення, тим менш впорядкованою і більш випадковою є система. Вчені називають міру випадковості або розладу в системі як ентропію. Висока ентропія означає високий розлад і низьку енергію. Молекули та хімічні реакції також мають різну ентропію. Наприклад, ентропія зростає, оскільки молекули при високій концентрації в одному місці дифундують і розтікаються. Другий закон термодинаміки говорить про те, що енергія завжди буде втрачатися як тепло при передачі енергії або перетвореннях.
Живі істоти високо впорядковані, вимагаючи постійного введення енергії, щоб підтримуватися в стані низької ентропії.
Потенціал і кінети
Коли об'єкт знаходиться в русі, є енергія, пов'язана з цим об'єктом. Подумайте про шкідливий м'яч. Навіть повільний шкідливий кулька може завдати великої шкоди іншим об'єктам. Енергія, пов'язана з об'єктами, що знаходяться в русі, називається кінетичною енергією (рис. 4.1.4). Швидкісна куля, людина, що йде, і швидкий рух молекул у повітрі (який виробляє тепло) мають кінетичну енергію.
А що робити, якщо той самий нерухомий кулька-шкідник піднятий на дві поверхи над землею краном? Якщо підвішений шкідливий куля нерухомий, чи є енергія, пов'язана з ним? Відповідь - так. Енергія, яка була потрібна для підняття шкідливого кулі, не зникла, а тепер зберігається в шкідливому кулі в силу свого положення і сили тяжіння, що діє на нього. Цей тип енергії називається потенційною енергією (рис. 4.1.4). Якби куля впала, потенційна енергія трансформувалася б в кінетичну енергію, поки вся потенційна енергія не вичерпається, коли м'яч спирався на землю. Шкідливі кулі також гойдаються як маятник; через гойдалки відбувається постійна зміна потенційної енергії (найвищий у верхній частині гойдалки) до кінетичної енергії (найвищий в нижній частині гойдалки). Інші приклади потенційної енергії включають енергію води, що утримується за греблею або людина збирається стрибати з парашутом з літака.
Потенційна енергія пов'язана не тільки з розташуванням матерії, але і зі структурою речовини. Навіть пружина на землі має потенційну енергію, якщо вона стискається; так само як і гумка, яка натягнута. На молекулярному рівні зв'язки, які утримують атоми молекул разом, існують в певній структурі, яка має потенційну енергію. Пам'ятайте, що анаболічні клітинні шляхи вимагають енергії для синтезу складних молекул з більш простих, а катаболічні шляхи вивільняють енергію, коли складні молекули розщеплюються. Той факт, що енергія може вивільнятися при розриві певних хімічних зв'язків, означає, що ці зв'язки мають потенційну енергію. Фактично, є потенційна енергія, що зберігається в зв'язках всіх молекул їжі, які ми їмо, яка в кінцевому підсумку використовується для використання. Це тому, що ці зв'язки можуть виділяти енергію при розриві. Тип потенційної енергії, яка існує в хімічних зв'язках і виділяється при розриві цих зв'язків, називається хімічною енергією. Хімічна енергія відповідає за забезпечення живих клітин енергією з їжі. Вивільнення енергії відбувається при порушенні молекулярних зв'язків всередині молекул їжі.
ПОНЯТТЯ В ДІЇ
Відвідайте сайт і виберіть «Маятник» в меню «Робота і енергія», щоб побачити зміщення кінетичної і потенційної енергії маятника в русі.
Вільна енергія та активація
Дізнавшись, що хімічні реакції вивільняють енергію при розриві зв'язків, що зберігають енергію, важливим наступним питанням є наступне: Як кількісно оцінюється та виражається енергія, пов'язана з цими хімічними реакціями? Як можна порівняти енергію, що виділяється від однієї реакції, з реакцією іншої? Вимірювання вільної енергії використовується для кількісної оцінки цих передач енергії. Нагадаємо, що згідно з другим законом термодинаміки, всі передачі енергії пов'язані з втратою деякої кількості енергії в непридатному вигляді, наприклад, тепла. Вільна енергія конкретно відноситься до енергії, пов'язаної з хімічною реакцією, яка доступна після врахування втрат. Іншими словами, вільна енергія - це корисна енергія, або енергія, яка доступна для виконання роботи.
Якщо енергія вивільняється під час хімічної реакції, то зміна вільної енергії, позначена як ∆G (дельта G), буде негативним числом. Негативна зміна вільної енергії також означає, що продукти реакції мають менше вільної енергії, ніж реагенти, тому що вони виділяють деяку кількість вільної енергії під час реакції. Реакції, які мають негативну зміну вільної енергії і, отже, вивільняють вільну енергію, називаються ексергоническими реакціями. Подумайте: exergonic означає, що енергія є колишньоюсистемою. Ці реакції також називають спонтанними реакціями, і їх продукти мають менше накопиченої енергії, ніж реагенти. Необхідно провести важливу відмінність між терміном спонтанний та ідеєю хімічної реакції, що відбувається негайно. Всупереч повсякденному вживанню терміна, спонтанна реакція - це не та, яка раптово або швидко виникає. Іржавіння заліза - приклад спонтанної реакції, яка відбувається повільно, потроху, з часом.
Якщо хімічна реакція поглинає енергію, а не вивільняє енергію на баланс, то ∆G для цієї реакції буде позитивним значенням. При цьому продукти мають більше вільної енергії, ніж реагенти. Таким чином, продукти цих реакцій можна розглядати як енергозберігаючі молекули. Ці хімічні реакції називаються ендергонічними реакціями і вони неспонтанні. Ендергонічна реакція не пройде сама по собі без додавання вільної енергії.
АРТ КОНК
Подивіться на кожен із показаних процесів і вирішіть, чи є він ендергонічним або ексергонічним.
Існує ще одна важлива концепція, яку необхідно враховувати щодо ендергонічних та ексергонічних реакцій. Ексергонічні реакції вимагають невеликої кількості енергії, щоб розпочати роботу, перш ніж вони зможуть продовжити свої кроки, що вивільняють енергію. Ці реакції мають чистий викид енергії, але все ж вимагають деякого введення енергії на початку. Ця невелика кількість вхідної енергії, необхідної для всіх хімічних реакцій, називається енергією активації.

Ферменти
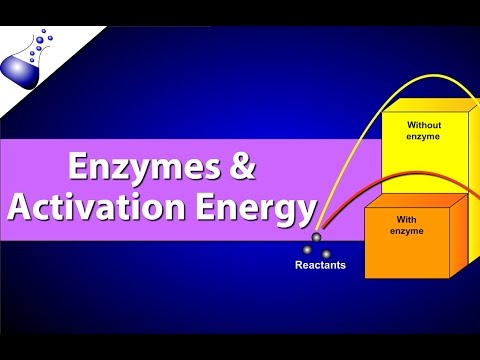
Речовина, яка допомагає протікати хімічній реакції, називається каталізатором, а молекули, що каталізують біохімічні реакції, називаються ферментами. Більшість ферментів є білками і виконують критичну задачу зниження енергій активації хімічних реакцій всередині клітини. Більшість реакцій, критичних для живої клітини, відбуваються занадто повільно при нормальних температурах, щоб бути корисним для клітини. Без ферментів для прискорення цих реакцій життя не могла б зберігатися. Ферменти роблять це, зв'язуючись з молекулами реагентів і утримуючи їх таким чином, щоб процеси хімічного розриву та формування зв'язків відбувалися легше. Важливо пам'ятати, що ферменти не змінюють, чи є реакція ексергонічною (спонтанною) або ендергонічною. Це тому, що вони не змінюють вільну енергію реагентів або продуктів. Вони лише зменшують енергію активації, необхідну для того, щоб реакція йшла вперед (рис. 4.1.6). Крім того, сам фермент незмінний реакцією, яку він каталізує. Після того, як одна реакція була каталізована, фермент здатний брати участь в інших реакціях.
Хімічні реагенти, з якими зв'язується фермент, називаються субстратами ферменту. Там може бути один або кілька субстратів, в залежності від конкретної хімічної реакції. У деяких реакціях один субстрат реагенту розбивається на кілька продуктів. В інших два субстрати можуть зібратися разом, щоб створити одну більшу молекулу. Два реагенти також можуть вступити в реакцію, і обидва модифікуються, але вони залишають реакцію як два продукти. Розташування всередині ферменту, де субстрат зв'язується, називається активним сайтом ферменту. Активний сайт - це місце, де відбувається «дія». Оскільки ферменти є білками, в активному місці існує унікальна комбінація бічних ланцюгів амінокислот. Кожна бічна ланцюг характеризується різними властивостями. Вони можуть бути великими або дрібними, слабокислотними або основними, гідрофільними або гідрофобними, позитивно або негативно зарядженими, або нейтральними. Унікальне поєднання бічних ланцюгів створює дуже специфічне хімічне середовище в межах активної ділянки. Це специфічне середовище підходить для зв'язування з одним конкретним хімічним субстратом (або субстратами).
Активні ділянки схильні до впливів місцевого середовища. Підвищення температури навколишнього середовища, як правило, збільшує швидкість реакції, каталізується ферментом або іншим чином. Однак температури поза оптимальним діапазоном знижують швидкість, з якою фермент каталізує реакцію. Гарячі температури в кінцевому підсумку спричинять денатуру ферментів, незворотну зміну тривимірної форми і, отже, функції ферменту. Ферменти також підходять для найкращого функціонування в межах певного діапазону концентрації рН та солі, і, як і при температурі, екстремальні концентрації рН та солі можуть призвести до денатурування ферментів.
Протягом багатьох років вчені думали, що зв'язування ферментів - субстрат відбувається простим способом «замок і ключ». Ця модель стверджувала, що фермент і субстрат ідеально поєднуються один з одним миттєвим кроком. Однак сучасні дослідження підтримують модель, яка називається індукованою посадкою (рис. 4.1.7). Модель з індукційним пристосуванням розширюється на моделі блокування та ключа, описуючи більш динамічне зв'язування між ферментом та субстратом. Коли фермент і субстрат об'єднуються, їх взаємодія викликає м'який зсув структури ферменту, який утворює ідеальний зв'язок між ферментом і субстратом.
Коли фермент пов'язує свій субстрат, утворюється ферментно-субстратний комплекс. Цей комплекс знижує енергію активації реакції і сприяє її швидкому прогресуванню одним з декількох можливих способів. На базовому рівні ферменти сприяють хімічним реакціям, які включають більше одного субстрату, об'єднуючи субстрати в оптимальній для реакції орієнтації. Інший спосіб, за допомогою якого ферменти сприяють реакції їх субстратів, - це створення оптимального середовища в межах активного місця для протікання реакції. Хімічні властивості, що випливають із конкретного розташування амінокислотних R груп в активному місці, створюють ідеальне середовище для реакції конкретних субстратів ферменту.
Комплекс фермент-субстрат також може знизити енергію активації, порушуючи структуру зв'язку, щоб її було легше розірвати. Нарешті, ферменти також можуть знижувати енергії активації, беручи участь у самій хімічній реакції. У цих випадках важливо пам'ятати, що фермент завжди повернеться в початковий стан до завершення реакції. Одним з характерних властивостей ферментів є те, що вони залишаються в кінцевому підсумку незмінними реакціями, які вони каталізують. Після того, як фермент каталізував реакцію, він вивільняє свій продукт (и) і може каталізувати нову реакцію.
Здавалося б, ідеальним є сценарій, при якому всі ферменти організму існували в рясному постачанні і функціонували оптимально за всіх клітинних умов, у всіх клітинках, у всі часи. Однак різноманітність механізмів гарантує, що цього не станеться. Клітинні потреби та умови постійно змінюються від клітини до клітини та змінюються в межах окремих клітин з часом. Необхідні ферменти клітин шлунка відрізняються від клітин зберігання жиру, клітин шкіри, клітин крові та нервових клітин. Крім того, клітина травного органу працює набагато важче, щоб переробляти та розщеплювати поживні речовини протягом часу, який уважно стежить за їжею порівняно з багатьма годинами після їжі. Оскільки ці клітинні потреби та умови змінюються, так і кількість та функціональність різних ферментів.
Оскільки швидкості біохімічних реакцій контролюються енергією активації, а ферменти знижуються і визначають енергію активації хімічних реакцій, відносні кількості і функціонування різноманітності ферментів всередині клітини в кінцевому підсумку визначають, які реакції будуть протікати і з якими темпами. Це визначення жорстко контролюється в осередках. У певних клітинних середовищах активність ферментів частково контролюється факторами навколишнього середовища, такими як рН, температура, концентрація солі та, в деяких випадках, кофактори або коферменти.
Ферменти також можна регулювати способами, які або сприяють, або зменшують активність ферментів. Існує багато видів молекул, які пригнічують або сприяють ферментативній функції, і різні механізми, за допомогою яких вони це роблять. У деяких випадках інгібування ферменту молекула інгібітора досить схожа на субстрат, який може зв'язуватися з активним ділянкою і просто блокувати субстрат від зв'язування. Коли це відбувається, фермент пригнічується за допомогою конкурентного інгібування, оскільки молекула інгібітора конкурує з субстратом за зв'язування з активним ділянкою.
З іншого боку, при неконкурентному інгібуванні молекула інгібітора зв'язується з ферментом у місці, відмінному від активного сайту, званому аллостеричним сайтом, але все ж вдається блокувати зв'язування субстрату з активним ділянкою. Деякі молекули інгібіторів зв'язуються з ферментами в місці, де їх зв'язування індукує конформаційне зміна, що знижує спорідненість ферменту до його субстрату. Цей тип гальмування називається аллостерическим інгібуванням (рис. 4.1.8). Більшість аллостерично регульованих ферментів складаються з більш ніж одного поліпептиду, що означає, що вони мають більше однієї білкової субодиниці. Коли аллостеричний інгібітор зв'язується з областю на ферменті, всі активні ділянки на білкових субодиницях дещо змінюються таким чином, що вони зв'язують свої субстрати з меншою ефективністю. Існують аллостеричні активатори, а також інгібітори. Алостеричні активатори зв'язуються з місцями на ферменті далеко від активного сайту, індукуючи конформаційні зміни, що підвищує спорідненість активного сайту (ів) ферменту для його субстрату (рис) (рис 4.1.8).
КАР'ЄРА В ДІЇ: Розробник фармацевтичних наркотиків
Ферменти є ключовими компонентами метаболічних шляхів. Розуміння того, як працюють ферменти та як їх можна регулювати, є ключовими принципами розробки багатьох фармацевтичних препаратів на сучасному ринку. Біологи, що працюють в цій галузі, співпрацюють з іншими вченими для розробки лікарських засобів (рис. 4.1.9).
Розглянемо статини, наприклад, статини - це назва одного класу препаратів, які можуть знизити рівень холестерину. Ці сполуки є інгібіторами ферменту ГМГ-КоА-редуктази, який є ферментом, що синтезує холестерин з ліпідів в організмі. Пригнічуючи цей фермент, рівень холестерину, що синтезується в організмі, може бути знижений. Аналогічно, ацетамінофен, в народі продається під торговою маркою Tylenol, є інгібітором ферменту циклооксигенази. Хоча він використовується для полегшення лихоманки та запалення (болю), його механізм дії досі не повністю вивчений.
Як виявляються наркотики? Однією з найбільших проблем у виявленні наркотиків є визначення мішені наркотиків. Мішень препарату - це молекула, яка буквально є мішенню препарату. У разі статинів мішенню препарату є ГМГ-КоА-редуктаза. Лікарські мішені виявляються шляхом кропітких досліджень в лабораторії. Ідентифікації однієї цілі недостатньо; вчені також повинні знати, як мішень діє всередині клітини і які реакції йдуть шкереберть у разі захворювання. Після того, як ціль і шлях ідентифіковані, то починається фактичний процес розробки препарату. На цьому етапі хіміки та біологи працюють разом, щоб розробити та синтезувати молекули, які можуть блокувати або активувати певну реакцію. Однак це лише початок: якщо і коли прототип препарату успішно виконує свою функцію, то він піддається багатьом тестам від експериментів in vitro до клінічних випробувань, перш ніж він зможе отримати схвалення від Управління з контролю за продуктами та ліками США, щоб бути на ринку.
Багато ферментів не працюють оптимально, або навіть взагалі, якщо вони не пов'язані з іншими специфічними небілковими молекулами-помічниками. Вони можуть тимчасово зв'язуватися через іонні або водневі зв'язки, або постійно через більш міцні ковалентні зв'язки. Зв'язування з цими молекулами сприяє оптимальній формі і функції їх відповідних ферментів. Два приклади цих типів молекул помічників - кофактори та коферменти. Кофакторами є неорганічні іони, такі як іони заліза і магнію. Коферменти - це органічні молекули помічників, ті, що мають основну атомну структуру, що складається з вуглецю та водню. Як і ферменти, ці молекули беруть участь у реакціях, не змінюючись самі, і в кінцевому підсумку переробляються та використовуються повторно. Вітаміни є джерелом коферментів. Одні вітаміни є попередниками коферментів, а інші діють безпосередньо як коферменти. Вітамін С є прямим коферментом для безлічі ферментів, які беруть участь в побудові важливої сполучної тканини, колагену. Тому ферментна функція частково регулюється великою кількістю різних кофакторів і коферментів, які можуть поставлятися дієтою організму або, в деяких випадках, вироблятися організмом.
Зворотній зв'язок гальмування в метаболічних шляхах
Молекули можуть регулювати функцію ферментів різними способами. Однак головне питання: що це за молекули і звідки вони беруться? Деякі з них є кофакторами та коферментами, як ви дізналися. Які ще молекули в клітині забезпечують ферментативну регуляцію, таку як аллостерична модуляція, а також конкурентне та неконкурентне гальмування? Мабуть, найбільш актуальними джерелами регуляторних молекул, щодо ферментативного клітинного метаболізму, є продукти самих клітинних метаболічних реакцій. Найбільш ефективним і елегантним способом клітини еволюціонували, щоб використовувати продукти власних реакцій для зворотного гальмування активності ферментів. Гальмування зворотного зв'язку передбачає використання продукту реакції для регулювання власного подальшого виробництва (рис. 4.1.10). Клітка реагує на велику кількість продуктів, сповільнюючи виробництво під час анаболічних або катаболічних реакцій. Такі продукти реакції можуть пригнічувати ферменти, які каталізували їх вироблення за допомогою описаних вище механізмів.
Вироблення як амінокислот, так і нуклеотидів контролюється за допомогою гальмування зворотного зв'язку. Додатково АТФ є аллостеріческим регулятором деяких ферментів, що беруть участь в катаболическом розпаді цукру, процесі, який створює АТФ. Таким чином, коли АТФ знаходиться в рясному постачанні, клітина може перешкоджати виробленню АТФ. З іншого боку, АДФ служить позитивним аллостеричним регулятором (аллостеричним активатором) для деяких тих же ферментів, які інгібуються АТФ. Таким чином, коли відносні рівні АДФ високі порівняно з АТФ, клітина спрацьовує виробляти більше АТФ через катаболізм цукру.
Підсумок розділу
Клітини виконують функції життєдіяльності за допомогою різних хімічних реакцій. Метаболізм клітини відноситься до поєднання хімічних реакцій, які відбуваються всередині неї. Катаболічні реакції розщеплюють складні хімічні речовини на більш прості і пов'язані з виділенням енергії. Анаболічні процеси будують складні молекули з більш простих і вимагають енергії.
Вивчаючи енергію, термін система позначає речовину та навколишнє середовище, що беруть участь у передачі енергії. Ентропія - це міра розладу системи. Фізичні закони, що описують передачу енергії, є законами термодинаміки. Перший закон говорить, що загальна кількість енергії у Всесвіті є постійним. Другий закон термодинаміки говорить, що кожна передача енергії передбачає деяку втрату енергії в непридатному вигляді, наприклад, теплової енергії. Енергія надходить у різних формах: кінетичній, потенційній та вільній. Зміна вільної енергії реакції може бути негативним (вивільняє енергію, ексергонічним) або позитивним (споживає енергію, ендергонічним). Всі реакції вимагають початкового введення енергії для продовження, званої енергією активації.
Ферменти є хімічними каталізаторами, які прискорюють хімічні реакції, знижуючи їх енергію активації. Ферменти мають активний сайт з унікальним хімічним середовищем, яке відповідає певним хімічним реагентам для цього ферменту, який називається субстратами. Вважається, що ферменти та субстрати зв'язуються відповідно до індукованої моделі. Дія ферменту регулюється для збереження ресурсів та оптимальної реакції на навколишнє середовище.
Мистецькі зв'язки
Малюнок 4.1.5: Подивіться на кожен із показаних процесів і вирішіть, чи є він ендергонічним або ексергонічним.
- Відповідь
-
Розкладання компостної купи - це ексергонічний процес. Малюк, що розвивається з заплідненої яйцеклітини, - це ендергонічний процес. Розчинення чаю у воді - це ексергонічний процес. Куля, що котиться вниз, - це ексергонічний процес.
Глосарій
- активація енергії
- кількість вихідної енергії, необхідної для протікання реакцій
- активний сайт
- конкретні області на фермент, де субстрат зв'язується
- аллостеричне гальмування
- механізм пригнічення дії ферменту, при якому регуляторна молекула зв'язується з другим ділянкою (не активним ділянкою) і ініціює зміну конформації в активному місці, запобігаючи зв'язування з субстратом
- анаболічні
- описує шлях, який вимагає чистої енергії для синтезу складних молекул з більш простих
- біоенергетика
- поняття потоку енергії через живі системи
- катаболічних
- описує шлях, в якому складні молекули розбиваються на більш прості, отримуючи енергію як додатковий продукт реакції
- конкурентне гальмування
- загальний механізм регулювання активності ферментів, в якому молекула, відмінна від субстрату ферменту, здатна зв'язувати активний сайт і перешкоджати зв'язуванню самого субстрату, тим самим пригнічуючи загальну швидкість реакції ферменту
- ендергонічні
- описує хімічну реакцію, яка призводить до продуктів, які зберігають більше хімічної потенційної енергії, ніж реагенти
- фермент
- молекули, що каталізує біохімічні реакції
- ексергонічних
- описує хімічну реакцію, яка призводить до продуктів з меншим хімічним потенціалом енергії, ніж реагенти, плюс вивільнення вільної енергії
- гальмування зворотного зв'язку
- механізм регулювання активності ферментів, в якому продукт реакції або кінцевий продукт серії послідовних реакцій інгібує фермент для більш ранньої стадії в серії реакцій
- теплова енергія
- енергія, передана від однієї системи до іншої, яка не працює
- кінетична енергія
- тип енергії, пов'язаної з об'єктами в русі
- метаболізм
- всі хімічні реакції, що протікають всередині клітин, в тому числі ті, які використовують енергію і ті, що вивільняють енергію
- неконкурентне гальмування
- загальний механізм регуляції активності ферментів, при якому регуляторна молекула зв'язується з ділянкою, відмінною від активного сайту, і не дає активному ділянці зв'язувати субстрат; таким чином, молекула інгібітора не конкурує з субстратом за активну ділянку; аллостерическое інгібування - це форма неконкурентне гальмування
- потенційна енергія
- тип енергії, яка відноситься до потенціал, щоб зробити роботу
- субстрат
- молекули, на яких діє фермент
- термодинаміка
- наука про взаємозв'язок між теплом, енергією та роботою