46.2: Потік енергії через екосистеми
- Page ID
- 1844

Цілі навчання
- Опишіть, як організми набувають енергію в харчовій павутині та в пов'язаних харчових ланцюгах
- Поясніть, як ефективність передачі енергії між трофічними рівнями впливає на структуру та динаміку екосистеми
- Обговоріть трофічні рівні та те, як екологічні піраміди використовуються для їх моделювання
Все живе вимагає енергії в тій чи іншій формі. Енергія потрібна більшості складних метаболічних шляхів (часто у вигляді аденозинтрифосфату, АТФ), особливо відповідальних за побудову великих молекул з менших сполук, а саме життя є енергетичним процесом. Живі організми не змогли б зібрати макромолекули (білки, ліпіди, нуклеїнові кислоти та складні вуглеводи) зі своїх мономерних субодиниць без постійного надходження енергії.
Важливо розуміти, як організми набувають енергію і як ця енергія передається від одного організму до іншого через харчові мережі та їх складові харчові ланцюги. Харчові мережі ілюструють, як енергія тече спрямовано через екосистеми, включаючи те, наскільки ефективно організми її набувають, використовують і скільки залишається для використання іншими організмами харчової павутини.
Як організми набувають енергії в харчовій павутині
Енергія набувається живими істотами трьома способами: фотосинтезом, хемосинтезом і споживанням і перетравленням інших живих або раніше живих організмів гетеротрофами.
Фотосинтетичні та хемосинтетичні організми об'єднуються в категорію, відому як автотрофи: організми, здатні синтезувати власну їжу (точніше, здатні використовувати неорганічний вуглець як джерело вуглецю). Фотосинтетичні автотрофи (фотоавтотрофи) використовують сонячне світло як джерело енергії, тоді як хемосинтетичні автотрофи (хемоавтотрофи) використовують неорганічні молекули як джерело енергії. Автотрофи мають вирішальне значення для всіх екосистем. Без цих організмів енергія була б недоступна іншим живим організмам і саме життя було б неможливим.
Фотоавтотрофи, такі як рослини, водорості та фотосинтезуючі бактерії, служать джерелом енергії для більшості екосистем світу. Ці екосистеми часто описуються випасом харчових павутин. Фотоавтотрофи використовують сонячну енергію сонця, перетворюючи її в хімічну енергію у вигляді АТФ (і НАДФ). Енергія, що зберігається в АТФ, використовується для синтезу складних органічних молекул, таких як глюкоза.
Хемоавтотрофи - це перш за все бактерії, які зустрічаються в рідкісних екосистемах, де сонячне світло недоступний, наприклад, у тих, що пов'язані з темними печерами або гідротермальними жерлами на дні океану (рис.\(\PageIndex{1}\)). Багато хемоавтотрофи в гідротермальних отворах використовують сірководень (H 2 S), який виділяється з вентиляційних отворів як джерело хімічної енергії. Це дозволяє хемоавтотрофам синтезувати складні органічні молекули, такі як глюкоза, для власної енергії і, в свою чергу, постачає енергію решті екосистеми.

Продуктивність в межах трофічних рівнів
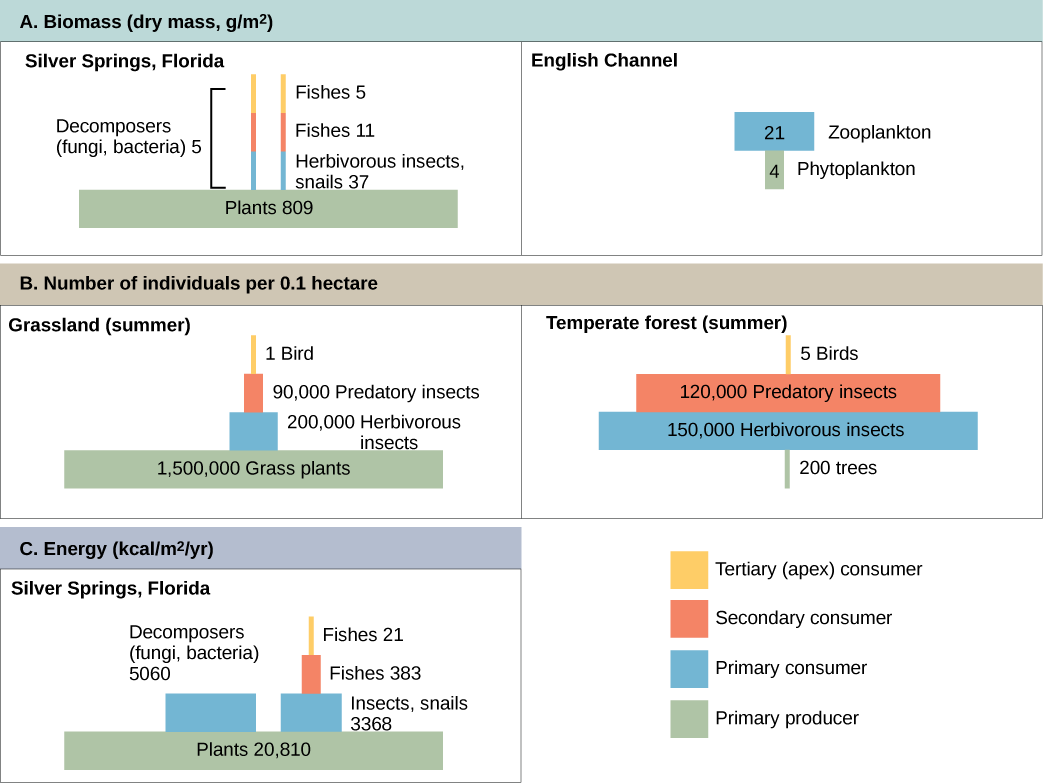
Продуктивність всередині екосистеми можна визначити як відсоток енергії, що надходить до екосистеми, включеної в біомасу на певному трофічному рівні. Біомаса - це загальна маса в одиниці площі на момент вимірювання живих або раніше живих організмів в межах трофічного рівня. Екосистеми мають характерну кількість біомаси на кожному трофічному рівні. Наприклад, в екосистемі Ла-Манш первинні виробники припадають на біомасу 4 г/м 2 (грам на метр у квадраті), тоді як основні споживачі демонструють біомасу 21 г/м 2.
Продуктивність первинних виробників особливо важлива в будь-якій екосистемі, оскільки ці організми приносять енергію іншим живим організмам шляхом фотоавтотрофії або хемоавтотрофії. Швидкість, з якою фотосинтетичні первинні виробники включають енергію від сонця, називається валовою первинною продуктивністю. Приклад валової первинної продуктивності показаний на блоковій діаграмі потоку енергії в водній екосистемі Silver Springs, як показано на малюнку 46.1.7. У цій екосистемі загальна енергія, накопичена первинними виробниками (валова первинна продуктивність), була показана 20,810 ккал/м 2 /рік.
Оскільки всім організмам потрібно використовувати частину цієї енергії для власних функцій (наприклад, дихання та метаболічних втрат тепла), вчені часто посилаються на чисту первинну продуктивність екосистеми. Чиста первинна продуктивність — це енергія, яка залишається у первинних виробників після обліку дихання та втрат тепла організмів. Потім чиста продуктивність доступна первинним споживачам на наступному трофічному рівні. У нашому прикладі Silver Spring 13 187 з 20,810 ккал/м 2/рік були використані для дихання або були втрачені як тепло, залишаючи 7,632 ккал/м 2/рік енергії для використання основними споживачами.
Екологічна ефективність: перенесення енергії між трофічними рівнями
Як показано на малюнку 46.1.7, велика кількість енергії втрачається з екосистеми від одного трофічного рівня до наступного рівня, оскільки енергія витікає від первинних виробників через різні трофічні рівні споживачів та розкладників. Основною причиною такої втрати є другий закон термодинаміки, який стверджує, що всякий раз, коли енергія перетворюється з однієї форми в іншу, в системі спостерігається тенденція до розладу (ентропії). У біологічних системах це означає, що багато енергії втрачається як метаболічне тепло, коли організми з одного трофічного рівня споживають наступний рівень. У прикладі екосистеми Silver Springs (рис. 46.1.7) ми бачимо, що первинні споживачі виробляли 1103 ккал/м 2 /рік з 7618 ккал/м 2/рік енергії, доступної їм від первинних виробників. Вимірювання ефективності передачі енергії між двома послідовними трофічними рівнями називається ефективністю передачі трофічного рівня (TLTE) і визначається за формулою:
\[\text{TLTE} = \frac{\text{production at present trophic level}}{\text{production at previous trophic level}} * 100 \nonumber\]
У Silver Springs TLTE між першими двома трофічними рівнями становив приблизно 14,8 відсотка. Низька ефективність передачі енергії між трофічними рівнями зазвичай є основним фактором, який обмежує довжину харчових ланцюгів, що спостерігаються в харчовій павутині. Справа в тому, що після чотирьох-шести передач енергії залишається недостатньо енергії для підтримки іншого трофічного рівня. У прикладі озера Онтаріо, показаному на малюнку 46.1.5, між первинним виробником (зелені водорості) та споживачем верхівки (лосось Чинук) відбулися лише три передачі енергії.
Екологи мають багато різних методів вимірювання перенесення енергії всередині екосистем. Деякі передачі легше або складніше виміряти залежно від складності екосистеми та того, скільки доступу вчені мають спостерігати за екосистемою. Іншими словами, деякі екосистеми важче вивчати, ніж інші, і іноді доводиться оцінювати кількісну оцінку передачі енергії.
Іншим основним параметром, який є важливим для характеристики потоку енергії в екосистемі, є чиста ефективність виробництва. Чиста ефективність виробництва (NPE) дозволяє екологам кількісно оцінити, наскільки ефективно організми певного трофічного рівня включають енергію, яку вони отримують в біомасу; вона розраховується за такою формулою:
\[\text{NPE} = \frac{\text{net consumer productivity}}{\text{assimilation}} * 100 \nonumber\]
Чиста споживча продуктивність - це енерговміст, доступний організмам наступного трофічного рівня. Асиміляція - це біомаса (вміст енергії, що генерується на одиницю площі) теперішнього трофічного рівня після обліку енергії, втраченої внаслідок неповного прийому їжі, енергії, що використовується для дихання, та енергії, втраченої як відходи. Неповний прийом їжі відноситься до того, що деякі споживачі з'їдають тільки частину своєї їжі. Наприклад, коли лев вбиває антилопу, він з'їсть все, крім шкури і кісток. Леву не вистачає багатого енергією кісткового мозку всередині кістки, тому лев не використовує всі калорії, які може забезпечити його здобич.
Таким чином, NPE вимірює, наскільки ефективно кожен трофічний рівень використовує та включає енергію з їжі в біомасу, щоб забезпечити наступний трофічний рівень. Взагалі холоднокровні тварини (ектотерми), такі як безхребетні, риби, земноводні та плазуни, використовують менше енергії, яку вони отримують для дихання та тепла, ніж теплокровні тварини (ендотерми), такі як птахи та ссавці. Додаткове тепло, що генерується в ендотермах, хоча і є перевагою з точки зору активності цих організмів в більш холодних середовищах, є головним недоліком з точки зору NPE. Тому багатьом ендотермам доводиться харчуватися частіше, ніж ектотерми, щоб отримати енергію, необхідну для виживання. Взагалі, NPE для ектотерм на порядок (10х) вище, ніж для ендотерм. Наприклад, NPE для гусениці, що їсть листя, вимірюється на рівні 18 відсотків, тоді як NPE для білки, що їсть жолуді, може становити лише 1,6 відсотка.
Неефективність використання енергії теплокровними тваринами має широкі наслідки для світового продовольства. Широко визнано, що м'ясна промисловість використовує велику кількість сільськогосподарських культур для годування худоби, а оскільки NPE низький, значна частина енергії від кормів для тварин втрачається. Наприклад, це коштує близько 1 цента, щоб виробляти 1000 дієтичних калорій (ккал) кукурудзи або сої, але приблизно $0,19 для виробництва аналогічної кількості калорій вирощування великої рогатої худоби для споживання яловичини. Така ж енергоємність молока від великої рогатої худоби також коштує дорого, приблизно в 0,16 долара на 1000 ккал. Значна частина цієї різниці обумовлена низьким NPE великої рогатої худоби. Таким чином, у всьому світі спостерігається зростаючий рух, спрямований на сприяння споживанню нем'ясних та немолочних продуктів, щоб менше енергії витрачалося даремно на годування тварин для м'ясної промисловості.
Моделювання енергетичного потоку екосистем: Екологічні піраміди
Структуру екосистем можна візуалізувати за допомогою екологічних пірамід, які вперше були описані новаторськими дослідженнями Чарльза Елтона в 1920-х роках. Екологічні піраміди показують відносну кількість різних параметрів (таких як кількість організмів, енергія та біомаса) на трофічних рівнях.
Піраміди чисел можуть бути як вертикальними, так і перевернутими, в залежності від екосистеми. Як показано на малюнку\(\PageIndex{2}\), типові пасовища протягом літа мають базу багатьох рослин і кількість організмів зменшується на кожному трофічному рівні. Однак протягом літа в помірному лісі основа піраміди складається з небагатьох дерев в порівнянні з кількістю первинних споживачів, в основному комах. Оскільки дерева великі, вони мають велику здатність до фотосинтезу та домінують над іншими рослинами в цій екосистемі для отримання сонячного світла. Навіть у меншій кількості первинні виробники в лісах все ще здатні підтримувати інші трофічні рівні.
Іншим способом візуалізації структури екосистеми є піраміди біомаси. Ця піраміда вимірює кількість енергії, перетвореної в живу тканину на різних трофічних рівнях. Використовуючи приклад екосистеми Silver Springs, ці дані демонструють вертикальну піраміду з біомаси (рис.\(\PageIndex{2}\)), тоді як піраміда з прикладу Ла-Маншу перевернута. Рослини (первинні виробники) екосистеми Silver Springs складають великий відсоток знайденої там біомаси. Однак фітопланктон у прикладі Ла-Маншу становить менше біомаси, ніж основні споживачі, зоопланктон. Як і у випадку з перевернутими пірамідами чисел, ця перевернута піраміда пов'язана не з недостатньою продуктивністю у первинних виробників, а є результатом високої швидкості обороту фітопланктону. Фітопланктон швидко споживається первинними споживачами, таким чином, мінімізуючи їх біомасу в будь-який конкретний момент часу. Однак фітопланктон швидко розмножується, таким чином вони здатні підтримувати решту екосистеми.
Моделювання екосистеми пірамід також може бути використано для показу потоку енергії через трофічні рівні. Зверніть увагу, що ці цифри такі ж, як ті, що використовуються на схемі енергетичного відсіку на малюнку 46.1.7. Піраміди енергії завжди вертикальні, і екосистема без достатньої первинної продуктивності не може бути підтримана. Всі види екологічних пірамід корисні для характеристики структури екосистеми. Однак при вивченні енергетичного потоку через екосистему піраміди енергії є найбільш послідовними і репрезентативними моделями структури екосистеми (рис.\(\PageIndex{2}\)).
Вправа
Піраміди, що зображують кількість організмів або біомаси, можуть бути перевернутими, вертикально або навіть ромбоподібними. Енергетичні піраміди, однак, завжди вертикально. Чому?
- Відповідь
-
Піраміди організмів можуть бути перевернутими або ромбоподібними, оскільки великий організм, такий як дерево, може підтримувати багато менших організмів. Аналогічним чином, низька біомаса організмів може підтримувати більшу біомасу на наступному трофічному рівні, оскільки організми швидко розмножуються і, таким чином, забезпечують безперервне живлення. Енергетичні піраміди, однак, завжди повинні бути вертикально через закони термодинаміки. Перший закон термодинаміки стверджує, що енергія не може бути ні створена, ні зруйнована; таким чином, кожен трофічний рівень повинен набувати енергію з трофічного рівня нижче. Другий закон термодинаміки стверджує, що під час передачі енергії деяка енергія завжди втрачається у вигляді тепла; таким чином, менше енергії є на кожному більш високому трофічному рівні.
Наслідки харчових павутин: біологічне збільшення
Одним з найважливіших екологічних наслідків динаміки екосистем є біомагніфікація. Біомагніфікація - це зростаюча концентрація стійких, токсичних речовин в організмах на кожному трофічному рівні, від первинних виробників до верхівкових споживачів. Було показано, що багато речовин біоакумулюються, включаючи класичні дослідження з пестицидом d іхлор d іфеніл т рихлоретан (ДДТ), який був опублікований у бестселері 1960-х років «Silent Spring» Рейчел Карсон. ДДТ був широко використовуваним пестицидом до того, як стало відомо про його небезпеку. У деяких водних екосистемах організми з кожного трофічного рівня споживали багато організмів нижчого рівня, що призвело до збільшення ДДТ птахів (споживачів верхівки), які їли рибу. Таким чином, птахи накопичили достатню кількість ДДТ, щоб викликати крихкість в яєчній шкаралупі. Цей ефект збільшив розбиття яєць під час гніздування і було показано, що він має несприятливий вплив на ці популяції птахів. Використання ДДТ було заборонено в США в 1970-х роках.
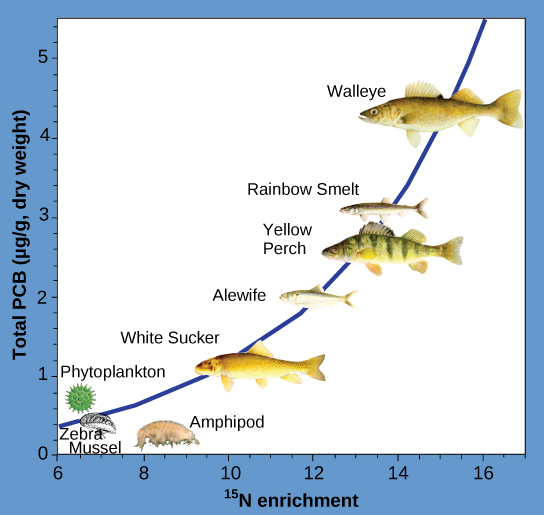
Інші речовини, які біозбільшують, - це поліхлоровані дифеніли (ПХБ), які використовувалися в рідинах-охолоджувачах у США, поки їх використання не було заборонено в 1979 році, та важкі метали, такі як ртуть, свинець та кадмій. Ці речовини найкраще вивчалися у водних екосистемах, де види риб на різних трофічних рівнях накопичують токсичні речовини, принесені через екосистему первинними виробниками. Як показано в дослідженні, проведеному Національним управлінням з питань океанічних та атмосферних впливів (NOAA) у затоці Сагіно озера Гурон (рис.\(\PageIndex{3}\)), концентрації друкованих плат збільшилися від первинних виробників екосистеми (фітопланктон) через різні трофічні рівні видів риб. Споживач верхівки (судака) має більш ніж у чотири рази більше ПХБ порівняно з фітопланктоном. Крім того, виходячи з результатів інших досліджень, птахи, які їдять ці риби, можуть мати рівні PCB принаймні на порядок вище, ніж ті, що містяться в озерній рибі.
Інші занепокоєння були викликані накопиченням важких металів, таких як ртуть та кадмій, у деяких видах морепродуктів. Агентство з охорони навколишнього середовища США (EPA) рекомендує вагітним жінкам та маленьким дітям не вживати жодної риби-меча, акули, королівської скумбрії або черепиці через високий вміст ртуті. Цим особам рекомендується їсти рибу з низьким вмістом ртуті: лосось, тілапія, креветки, минтай, сом. Біомагніфікація є хорошим прикладом того, як динаміка екосистем може впливати на наше повсякденне життя, навіть впливаючи на їжу, яку ми їмо.
Резюме
Організми в екосистемі отримують енергію різними способами, яка передається між трофічними рівнями, коли енергія тече знизу до верхньої частини харчової павутини, при цьому енергія втрачається при кожній передачі. Ефективність цих переносів важлива для розуміння різної поведінки та харчових звичок теплокровних проти холоднокровних тварин. Моделювання енергії екосистеми найкраще проводити за допомогою екологічних пірамід енергії, хоча інші екологічні піраміди надають іншу життєво важливу інформацію про структуру екосистеми.
Глосарій
- засвоєння
- біомаса, споживана і засвоюється з попереднього трофічного рівня після обліку енергії, втраченої внаслідок неповного прийому їжі, енергії, що використовується для дихання, і енергії, втраченої як відходи
- біомагніфікація
- підвищення концентрації стійких, токсичних речовин в організмах на кожному трофічному рівні, від первинних продуцентів до верхівкових споживачів
- біомаси
- загальна вага, на момент вимірювання, живих або раніше живих організмів в одиниці площі в межах трофічного рівня
- хемоавтотроф
- організм здатний синтезувати власну їжу, використовуючи енергію з неорганічних молекул
- екологічна піраміда
- (також, Ельтонівська піраміда) графічне зображення різних трофічних рівнів в екосистемі на основі кількості організму, біомаси або вмісту енергії
- валова первинна продуктивність
- швидкість, з якою фотосинтетичні первинні виробники включають енергію від сонця
- чиста споживча продуктивність
- енергетичний вміст, доступне організмам наступного трофічного рівня
- чиста первинна продуктивність
- енергія, яка залишається в первинних виробниках після обліку дихання і тепловтрат організмів
- чиста ефективність виробництва (NPE)
- міра здатності трофічного рівня перетворювати енергію, яку він отримує від попереднього трофічного рівня, в біомасу
- ефективність передачі трофічного рівня (TLTE)
- ефективність передачі енергії між двома послідовними трофічними рівнями