3.3: Ядро
- Page ID
- 5125

Ядро є відмінною рисою еукаріотичних клітин; сам термін еукаріот означає наявність «справжнього ядра».
Ядерна оболонка
Ядро обволікається парою мембран, що охоплюють просвіт, який є безперервним з просвітом ендоплазматичної сітки. Внутрішня мембрана стабілізується сіткою проміжних білків ниток, званих ламінами. Ядерна оболонка перфорована тисячами ядерних порних комплексів (NPC), які контролюють проходження молекул всередину і з ядра.
Хроматин
 Ядро містить хромосоми клітини. Кожна хромосома складається з однієї молекули ДНК, комплексної з рівною масою білків. У сукупності ДНК ядра з пов'язаними з ним білками називається хроматином.
Ядро містить хромосоми клітини. Кожна хромосома складається з однієї молекули ДНК, комплексної з рівною масою білків. У сукупності ДНК ядра з пов'язаними з ним білками називається хроматином.
Велика частина білка складається з декількох копій 5 видів гістонів. Це основні білки, щетинисті позитивно зарядженими залишками аргініну і лізину. (І Arg, і Lys мають вільну аміногрупу на своїй групі R, яка приваблює протони (H +), даючи їм позитивний заряд.) Просто вибір амінокислот, які ви зробили б, щоб щільно зв'язуватися з негативно зарядженими фосфатними групами ДНК.
Хроматин також містить невелику кількість найрізноманітніших негістонових білків. Більшість з них є факторами транскрипції (наприклад, стероїдних рецепторів), і їх зв'язок з ДНК є більш тимчасовим.
На зображенні праворуч показані 5 гістонів, розділених електрофорезом. Ці 5 білків мало відрізняються від одного типу клітин до іншого або навіть від одного виду до іншого. Однак багато негістонових білків у хроматіні (також показано праворуч) змінюються від одного типу клітин до іншого та від одного виду до іншого. (Надано Гері Стейн і Джанет Свінхарт Стайн, Університет Флориди.)
Дві копії кожного з чотирьох видів гістонів H2A, H2B, H3, H4 утворюють ядро білка, ядро нуклеосоми. Навколо цього загорнуто близько 147 пар основи ДНК. Від 20—60 б.п. ДНК пов'язують одну нуклеосому до наступної. Кожна область компонувальника зайнята однією молекулою гістону 1 (Н1). Ця область довша (50—150 б.п.) поруч з промоторами генів, що, імовірно, робить більше місця для зв'язування факторів транскрипції.
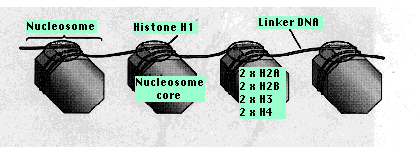
Нуклеосоми Схема нуклеосом
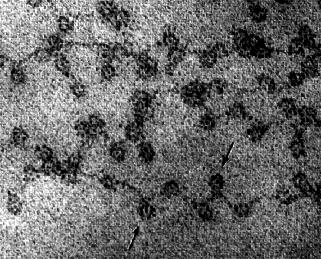
Зв'язування гістонів з ДНК не залежить від конкретних нуклеотидних послідовностей в ДНК, але критично залежить від амінокислотної послідовності гістону. Гістони - одні з найбільш збережених молекул в ході еволюції. Гістон Н4 у теляти відрізняється від Н4 в рослині гороху всього 2 залишками амінокислот в ланцюжку 102. На наведеній вище електронній мікрофотографії (люб'язно надано Девідом Е. Олінсом і Адою Л. Олінсом) для нуклеосом показано хроматин з ядра курячого еритроцита (птахи, на відміну від більшості ссавців, зберігають ядро в своїх зрілих еритроцитах). Стрілки вказують на нуклеосоми. Можна зрозуміти, чому розташування нуклеосом уподібнили «намистинам на нитці».
Освіта нуклеосом допомагає дещо, але майже не достатньо, зробити ДНК досить компактною, щоб поміститися в ядрі. Для того, щоб помістити 46 молекул ДНК (у людини), загальною довжиною понад 2 метри, в ядро, яке може бути лише 10 мкм в поперечнику, потрібно більш широке складання та ущільнення. Взаємодія між оголеними «хвостами» основних гістонів змушує нуклеосоми асоціюватися в компактне волокно діаметром 30 нм. Потім ці волокна складаються в більш складні структури, точна конфігурація яких невизначена і яка, ймовірно, змінюється з рівнем активності генів у регіоні.
Модифікації гістонів
Хоча їх амінокислотна послідовність (первинна структура) незмінна, окремі молекули гістону змінюються за структурою в результаті хімічних модифікацій, які відбуваються пізніше окремих амінокислот. До них відносяться додавання:
- ацетильні групи (CH 3 CO−) до лізинів
- фосфатні групи до серинів і треонінів
- метильні групи до лізинів і аргінінів
Хоча 75— 80% молекули гістону включені в ядро, решта - на N-терміналі - бовтається з ядра як «хвіст» (не показано на малюнку). Більшість хімічних модифікацій відбувається на цих хвостах, особливо H3 і H4. Більшість змін тез є оборотними. Наприклад, ацетильні групи додаються ферментами, званими гістоновими ацетилтрансферазами (HAT) (не плутати із середовищем «HAT», що використовується для виготовлення моноклональних антитіл), а також видаляються гістоновими деацетилазами (HDAC). Найчастіше ацетилювання гістонів відбувається в областях хроматину, які стають активними при транскрипції генів. Це робить свого роду інтуїтивний сенс, оскільки додавання ацетильних груп нейтралізує позитивні заряди на Lys, тим самим зменшуючи силу зв'язку між високонегативною ДНК та високопозитивними гістонами.
Однак, безумовно, є більше історії. Ацетилювання Lys-16 на H4 («H4K16aC») запобігає взаємодії їх «хвостів», необхідних для формування компактної 30-нм структури неактивного хроматину і, таким чином, асоціюється з активними генами (зверніть увагу, що цей випадок передбачає переривання білково-білкових, а не білково-ДНК взаємодій). Метилювання, яке також нейтралізує заряд на лізини (та аргініни), може або стимулювати, або пригнічувати транскрипцію генів у цій області.
- Додавання 3 метильних груп до лізин-4 та/або лізин-36 в H3 (H3K4Me3 та H3K36Me3 відповідно) пов'язане з активною транскрипцією генів, тоді як
- триметилювання лізину-9 та/або лізину-27 в H3 (H3k9Me3 і H3K27Me3 відповідно) пов'язане з неактивними генами. (До них відносяться ті відбиті гени, які були постійно інактивовані в соматичних клітині.)
- А додавання фосфатів призводить до того, що хромосоми стають більш - не менш компактними, оскільки вони готуються до мітозу та мейозу.
У будь-якому випадку тепер зрозуміло, що гістони є динамічною складовою хроматину, а не просто інертним ДНК-пакувальним матеріалом.
Варіанти гістонів
У нас є гени для 8 різних різновидів гістону 1 (H1). Яка різновид зустрічається у конкретного лінкера, залежить від таких факторів, як тип клітини, де вона знаходиться в клітинному циклі, і її стадія диференціації. У деяких випадках, принаймні, конкретний варіант H1 асоціюється з певними факторами транскрипції, щоб зв'язуватися з підсилювачем конкретних генів, вимикаючи експресію цих генів.
Деякі інші приклади варіантів гістонів:
- Н3 замінюється CENP-A («центромерний білок А») на нуклеосомах поблизу центромерів. Неможливість замінити CENP-A на Н3 в цих регіонах блокує структуру і функцію центромерів.
- H2A замінюється варіантом H2A.Z у генних промоторів і підсилювачів.
- Всі «стандартні» гістони замінюються варіантами в міру розвитку сперми.
Загалом, «стандартні» гістони включаються в нуклеосоми, оскільки нова ДНК синтезується під час S фази клітинного циклу. Пізніше деякі замінюються варіантними гістонами, як диктують умови в клітині.
Території хромосом
Під час інтерфази мало видно структури хроматину (за винятком особливих випадків, таких як політенні хромосоми дрозофіли та деяких інших мух). Хоча кожна хромосома сильно витягнута, вона, як правило, займає дискретну область в межах ядра, яка називається його територією. Це можна продемонструвати:
- направляючи крихітний лазерний промінь на невелику частину ядра. Якби всі хромосоми були переплетені, можна було б очікувати, що всі отримають деяку шкоду. Цього не відбувається — пошкоджується лише одна або дві хромосоми.
- Флуоресцентні плями, специфічні для конкретної хромосоми, забарвлюють лише дві області в ядрі — виявляючи територію двох гомологів.
«Цілуються» хромосоми
Частини однієї хромосоми можуть виходити з її території і взаємодіяти з частиною іншої хромосоми, що випливає з її території. Це «цілуються» хромосоми. Приклади, які були знайдені до цих пір, вказують на те, що ці взаємодії є ще одним способом координації активності генів, що мешкають на різних хромосомах.
Геном людини містить багато генів - розкиданих уздовж різних хромосом - які включаються при надходженні одного сигналу. Серед багатьох генів, активованих естрогеном, є TIFF1 на хромосомі 21 і GREB1 на хромосомі 2. Використовуючи аналіз FISH, дослідники Каліфорнійського університету в Сан-Дієго показали, що протягом лише 2 хвилин після впливу клітин естрогену локуси TIFF1 та GREB1 переміщаються з відповідних хромосомних територій та «поцілунок».
У миші, наївні помічники Т-клітин - очікують сигналу, щоб направити їх стати або Th1 клітинами, або Th2 клітинами
- частина хромосоми 10, що несе ген інтерферон-гамма (цитокіну Th1) цілування
- частина хромосоми 11, що несе гени для IL-4 і IL-5 (цитокінів Th2).
Коли клітина отримує сигнали, що здійснюють її на той чи інший шлях, дві області відокремлюються, відповідна йде в область активної транскрипції; інша - до області гетерохроматину.
І ще один приклад (в даному випадку два локуса далеко один від одного на тій же хромосомі «поцілунок»):
В області голови личинки дрозофіли припиняється експресія генів гомеобкса (HOX) Antp і Abd-B. Аналіз FISH показує, що ці два локуси - 10 000 000 пар основ окремо на хромосомі III - об'єднані в ядрі, пов'язаному білками, які перешкоджають їх транскрипції.
Еухроматин проти гетерохроматину
Щільність хроматину, що становить кожну хромосому (тобто наскільки щільно вона упакована) змінюється по довжині хромосоми. Щільні області називаються гетерохроматином, а менш щільні - евхроматином. Гетерохроматин знаходиться в частинок хромосоми, де мало або взагалі відсутні гени, такі як
- центромерів і
- теломери. Цей гетерохроматин міститься у всіх типах клітин організму.
- також зустрічається в багатих генами областях хромосоми, але там, де гени неактивні; тобто не транскрибується. Розташування цього гетерохроматину варіюється від одного типу диференційованої клітини до іншої (як ми очікували) — клітина печінки, наприклад, повинна припинити експресію генів, які не потрібні для її функцій.
- щільно упакований.
- значно збагачується транспозонами та іншими типами ДНК, які не сприяють протеому.
- реплікується пізно в S-фазі клітинного циклу.
- має зниження перетину при мейозі.
- локалізується поблизу внутрішньої поверхні ядерної оболонки, в більшості клітин тварин.
- Гістони в нуклеосомах гетерохроматину показують характерні модифікації:
- зниження ацетилювання;
- підвищене метилювання лізину-9 в гістоні H3 (H3K9), який тепер забезпечує місце зв'язування гетерохроматинового білка 1 (HP1), який блокує доступ факторами транскрипції, необхідними для транскрипції генів
- підвищення метилювання лізину-27 в гістоні H3 (H3K27)
Еухроматин
- зустрічається в частинок хромосоми, які містять багато активних генів.
- нещільно упакований в петлі з 30-нм волокон.
- відокремлені від сусіднього гетеро хроматину ізоляторами.
- У клітині тварин еухроматин і, таким чином, активна транскрипція генів відбувається поблизу центру ядра.
- Гени в еухроматині показують
- зниження метилювання цитозинів у ділянках CpG промотора (ів) гена
- підвищене ацетилювання прилеглих гістонів
- зниження метилювання лізину-9 та лізину-27 в гістоні H3
Нуклеосоми і транскрипція
Фактори транскрипції не можуть зв'язуватися зі своїм промотором, якщо промоутер заблокований нуклеосомою. Однією з перших функцій факторів транскрипції збірки є або вигнання нуклеосоми з місця, де починається транскрипція, або принаймні ковзати нуклеосоми вздовж молекули ДНК. Будь-яка дія викриває промотор гена, так що фактори транскрипції можуть потім зв'язуватися з ним.
Фактична транскрипція генів, що кодують білок, здійснюється РНК-полімеразою II (Pol II або RNAP II). Для того, щоб він подорожував по ДНК, щоб транскрибувати, комплекс білків видаляє нуклеосоми перед ним, а потім замінює їх після того, як Пол II транскрибує цю частину ДНК і рухається далі.
Нуклеосоми та реплікація ДНК
Як і у випадку з транскрипцією, спіраль ДНК повинна відкритися, щоб продовжити реплікацію ДНК. Це також вимагає, щоб нуклеосоми, що передують вилці реплікації, були видалені, а потім швидко зібрані, коли синтезуються провідні та відстаючі нитки.
Ядро
У період між клітинними поділами, коли хромосоми знаходяться в їх розширеному стані, одна або кілька з них (10 в клітині людини) мають петлі, що тягнуться в сферичну масу, звану ядерцем. Тут синтезуються три (з чотирьох) видів молекул РНК (28S, 18S, 5.8S), що використовуються при складанні великих і малих субодиниць рибосом.
28S, 18S та 5.8S рибосомальна РНК транскрибується (РНК-полімеразою I) від сотень до тисяч тандемічно розташованих генів рДНК, розподілених (у людини) на 10 різних хромосомах. РДНК-містять області цих 10 хромосом скупчуються в ядерці.
(У дріжджах молекули 5S рРНК - а також молекули переносної РНК - також синтезуються (РНК-полімеразою III) в ядерці.)
Після утворення молекули рРНК асоціюються з десятками різних рибосомних білків, що використовуються при складанні великих і малих субодиниць рибосоми.
Але білки синтезуються в цитозолі - і всі рибосоми необхідні в цитозолі для виконання своєї роботи - тому повинен бути механізм транспортування цих великих структур всередину і з ядра. Це одна з функцій ядерних порних комплексів.
Ядерні порні комплекси (NPC)
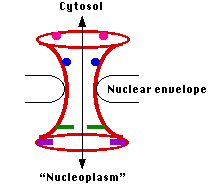 Ядерна оболонка перфорована тисячами пір. Кожен побудований з декількох копій декількох десятків різних білків, званих нуклеопоринами.
Ядерна оболонка перфорована тисячами пір. Кожен побудований з декількох копій декількох десятків різних білків, званих нуклеопоринами.
Весь вузол утворює водний канал, що з'єднує цитозол з внутрішньою частиною ядра («нуклеоплазма»). Коли матеріали повинні транспортуватися через пори, він відкривається, утворюючи канал шириною близько 27—41 нм - досить великий, щоб отримати такі великі збірки, як рибосомні субодиниці.
Транспорт через ядерні порні комплекси активний; тобто вимагає
- енергетика
- багато різних молекул носіїв, кожна спеціалізується на транспортуванні певного вантажу
- стикувальні молекули в НПК (представлені тут у вигляді кольорових стрижнів і дисків)
Імпорт в ядро
Білки синтезуються в цитозолі, і ті, які потрібні ядру, повинні бути імпортовані в нього через НПС. До них відносяться:
- всі гістони, необхідні для створення нуклеосом
- всі рибосомальні білки, необхідні для складання рибосом
- всі фактори транскрипції (наприклад, стероїдні рецептори), необхідні для включення генів (і виключення)
- всі фактори зрощування, необхідні для обробки попередньої мРНК у зрілі молекули мРНК; тобто для вирізання інтронних областей та зрощування екзонових областей.
Ймовірно, всі ці білки мають характерну послідовність амінокислот - називається послідовністю ядерної локалізації (NLS) - які націлені на їх введення.
Експорт з ядра
Молекули та високомолекулярні збірки, експортовані з ядра, включають:
- рибосомні субодиниці, що містять як рРНК, так і білки
- молекули месенджерної РНК (мРНК) (супроводжуються білками)
- переносять молекули РНК (тРНК) (також супроводжуються білками)
- фактори транскрипції, які повертаються до цитозолу, щоб чекати повторного використання
Як РНК, так і молекули білка містять характерну послідовність ядерного експорту (NES), необхідну для забезпечення їх зв'язку з правильними молекулами-носіями, щоб вивести їх у цитозол.