8.5: Рецепторні тирозинкінази (РТК)
- Page ID
- 4577

Рецепторні тирозинкінази опосередковують відповіді на велику кількість сигналів, включаючи пептидні гормони, такі як інсулін та фактори росту, такі як епідермальний фактор росту. Як і GPCR, рецепторні тирозинкінази зв'язують сигнал, потім передають повідомлення через ряд внутрішньоклітинних молекул, остання з яких діє на білки-мішені для зміни стану клітини.
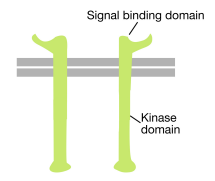
Як випливає з назви, рецептор тирозинкінази - це рецептор поверхні клітин, який також має активність тирозинкінази. Домен, що зв'язує сигнал тирозинкінази рецептора, знаходиться на поверхні клітини, в той час як ферментативна активність тирозинкінази знаходиться в цитоплазматичної частини білка (див. Малюнок вище). Трансмембранна альфа-спіраль з'єднує ці дві області рецептора.
Що відбувається, коли сигнальні молекули зв'язуються з рецепторними тирозинкіназами?
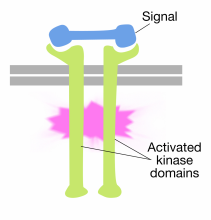
Зв'язування сигнальних молекул з позаклітинними доменами молекул рецепторної тирозинкінази змушує дві молекули рецепторів димеризуватися (об'єднуватися і асоціюватися). Це наближає цитоплазматичні хвости рецепторів близько один до одного і призводить до включення активності тирозинкіназ цих хвостів. Потім активовані хвости фосфориліруют один одного на кілька залишків тирозину. Це називається аутофосфорилирование.
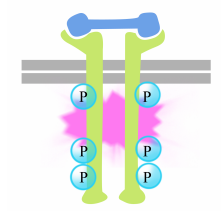
Фосфорилювання тирозинів на хвостах рецепторів запускає складання внутрішньоклітинного сигнального комплексу на хвостах. Нещодавно фосфорильовані тирозини служать сайтами зв'язування для сигналізації білків, які потім передають повідомлення ще іншим білкам. Важливий білок, який згодом активується сигнальними комплексами на рецепторних тирозинкіназах, називається Рас.
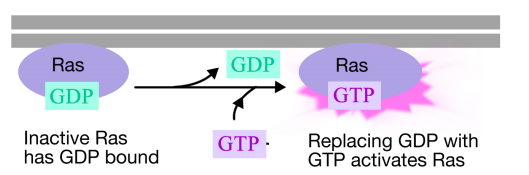
Білок Ras - це мономерний гуанін-нуклеотидний зв'язуючий білок, який пов'язаний з цитозольної поверхнею плазматичної мембрани (насправді він багато в чому схожий на альфа-субодиницю тримерних G-білків). Так само, як альфа-субодиниця G-білка, Ras активний, коли GTP пов'язаний з ним і неактивний, коли ВВП пов'язаний з ним. Також, як і альфа-субодиниця, Ras може гідролізувати GTP до ВВП.
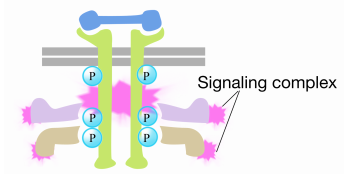
Коли сигнал надходить на рецепторну тирозинкіназу, мономери рецепторів об'єднуються і фосфорилюють тирозини один одного, запускаючи збірку комплексу білків на цитоплазматичному хвості рецептора. Один з білків в цьому комплексі взаємодіє з Рас і стимулює обмін ВВП, пов'язаного з неактивним Рас на ГТП. Це активує Рас.
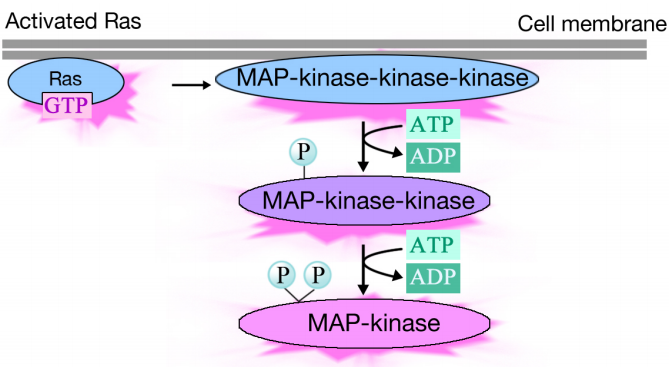
Активоване Ras запускає фосфориліруючий каскад з трьох протеїнкіназ, які ретранслюють і розподіляють сигнал. Ці білкові кінази входять до групи, яка називається MAP кінази (Mitogen Activated Proteint-kinases). Кінцева кіназа в цьому каскаді фосфорилює різні білки-мішені, включаючи ферменти і транскрипційні активатори, які регулюють експресію генів.
Фосфорилювання різних ферментів може змінювати їх діяльність та запускати нові хімічні реакції в клітині, тоді як фосфорилювання активаторів транскрипції може змінювати, які гени виражені. Комбінований ефект змін експресії генів та активності білка змінює фізіологічний стан клітини.
Знову ж таки, слідуючи шляху трансдукції сигналу, опосередкованого RTK, можна розрізнити ту саму основну картину подій: сигнал пов'язаний позаклітинними доменами рецепторних тирозинкіназ, що призводить до димеризації рецепторів та аутофосфорилювання цитозольних хвостів, таким чином передаючи повідомлення в інтер'єр клітини.
Повідомлення передається через сигнальний комплекс Рас, який потім стимулює серію кіназ. Термінальна кіназа в каскаді діє на білки-мішені і призводить до зміни білкової активності та експресії генів.
Описи вище дають дуже простий ескіз деяких основних класів рецепторів і стосуються в першу чергу механістичних деталей кроків, за допомогою яких сигнали, отримані різними типами рецепторів, викликають зміни в клітині. Основним заняттям додому є істотна схожість різних шляхів.
Ще один момент, який слід пам'ятати, полягає в тому, що, хоча ми розглядали кожен окремий шлях ізольовано, клітина в будь-який момент часу отримує кілька сигналів, які запускають різні відповіді одночасно. Описані вище шляхи показують значну ступінь «перехресного розмови», і на реакцію на будь-який заданий сигнал впливають інші сигнали, які клітина отримує одночасно. Безліч різних рецепторів, сигналів і їх комбінацій є засобами, за допомогою яких клітини здатні реагувати на величезну різноманітність різних обставин.