2.6: Лак Оперон; CAP сайт; ДНК-сліди
- Page ID
- 7557

Лак оперон кишкової палички
.png)
Малюнок 2.6.1: Лак Оперон
- Лак-Z коди для b -галактозидази, яка є ферментом, який розщеплює b -галактозиди (наприклад, лактозу).
- lac Y коди для пермеази, яка бере участь у транспортуванні b -галактозидів в клітину.
- Лак-А коди для b -галактозиду трансацетилази, яка ацетилює b -галактозиди.
- Мутація в лаку Z або лаку Y може призвести до лаку - генотипу, тобто клітин, які не можуть використовувати b -галактозиди як поживну речовину.
- Лак-А-мутант, який не має активності трансацетилази, все ще може утилізувати b -галактозиди (це все ще лак+ генотип). Його роль в метаболізмі b галактозидів не ясна.
Промоутер
область ДНК, яка бере участь у зв'язуванні РНК-полімерази для початку транскрипції.
Термінатор
послідовність ДНК, яка змушує РНК-полімеразу припиняти транскрипцію.
- Скупчення трьох генів, lac ZYA, транскрибується в одну мРНК (поліцистронічне повідомлення) від промотора тільки вище за течією від гена lac Z.
- При відсутності індуктора кластер генів не транскрибується.
- Коли індуктор додається (наприклад, лактоза, або негідролізуючий аналоговий ізопропілтіогалактозид - IPTG) транскрипція починається з одного промотора (лак Р) і проходить через гени лаку ZYA до послідовності термінатора, розташованої нижче за течією Лак А ген.
Примітка
Лак-ZYA мРНК має період напіврозпаду ~ 3 хвилини, що дозволяє індукції відносно швидко скасовуватися (тобто клітини припиняють виробляти ферменти швидко після зупинки індукції).
Вправа 2.6.1
З якою молекулою взаємодіє індуктор (лактоза), щоб впливати на транскрипційну регуляцію (тобто індукцію лак-оперону)?
- Відповідь
-
Це не b-галактозидаза, пермаза або трансацетилаза, скоріше це окремий білок, який називається білком репресора.
- Гени лаку контролюються механізмом, званим негативною регуляцією.
- Це означає, що вони транскрибуються, якщо вони не вимкнені білком регулятора.
- Мутація, яка інактивує білок регулятора, викликає постійну експресію генів lac ZYA.
Існує два типи генів в лак-опероні:
- Структурні гени - вони кодують ферменти, необхідні для деякого біохімічного шляху (наприклад, лак Z, Y і A).
- Гени-регулятори - вони кодують для білків, що беруть участь в регуляції структурних генів.
лак I - ген-регулятор лак-оперона.
- Цей ген розташований лише вище за течією області промотора для структурних генів лаку.
- Ген Lac I має свій промотор (конститутивний) і термінатор.
- Він робить моноцистроническое повідомлення, і кодує на один білок - білок лак-репресор.
- Це може запобігти транскрипції
- Він може розпізнавати та зв'язувати індуктор малих молекул (лактоза або IPTG)
Профілактика транскрипції лак-репресором
- лак-репресор (активний як тетрамерний білок) зв'язується з послідовністю ДНК, яка називається оператором (область лаку О).
- Область оператора лежить між областю промотора лаку (ділянкою зв'язування РНК-полімерази та ініціації транскрипції) та геном Lac Z.
- Перші 26 пар основ гена lac Z складають операторну область.
- Це не те, що протеїн-репресор «блокує» рух РНК-полімерази через ген lac Z.
- Репресорне зв'язування та зв'язування РНК-полімерази (з промотором) є взаємовиключними в області лак-промотора/оператора (lac PO).
Як змінюється взаємодія репресор/оператора в присутності молекули індуктора?
- Індуктор може зв'язуватися з репресором, утворюючи комплекс репресор/індуктор, який більше не асоціюється з оператором.
- Ключовою особливістю цієї взаємодії є те, що білок репресора має два місця зв'язування, один для індуктора і один для оператора.
- Коли індуктор зв'язується на своєму місці, він змінює конформацію білка репресора таким чином, що сайт зв'язування оператора має значно знижену спорідненість до області оператора ДНК.
- Такий вид контролю називається аллостеричним контролем.
- Результатом є те, що при додаванні індуктора репресор перетворюється в форму, яка звільняє від оператора.
.png)
Малюнок 2.6.2: Індуктор
Позитивний контроль лак-оперона здійснює комплекс Camp-cap
- кишкова паличка воліє глюкозу перед іншими джерелами вуглецю.
- Коли кишкова паличка вирощується на глюкозі, якщо додається інший цукор (наприклад, лактоза), індукція ферментів для утилізації іншого цукру не відбувається, поки глюкоза не буде витрачена.
- Коли кишкова паличка голодує за глюкозою, вона синтезує незвичайний нуклеотид: циклічний 3'5' аденозинмонофосфат (циклічний AMP, або цАМФ):
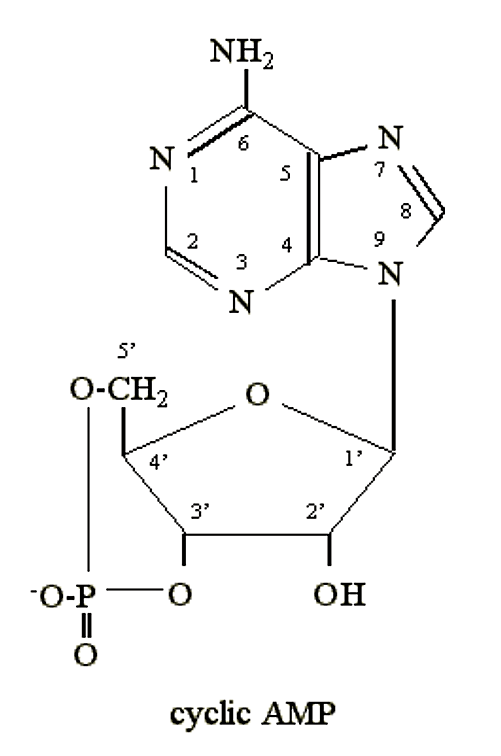.png)
Малюнок 2.6.3: цАМФ
- У бактерій підвищення рівня цАМФ здається «тривожним» сигналом, що вказує на низький рівень глюкози:
.png)
Малюнок 2.6.4: Взаємодія рівня цАМФ та лак-оперона
Дибутирил цАМФ
- аналог цАМФ, який може проходити через мембрану кишкової палички і в клітину.
- Якщо це додати до середовищ, що містять глюкозу і лактозу, це призведе до індукції лак-оперона.
- Таким чином, він імітує хімічне повідомлення, яке обманює кишкову паличку реагувати так, ніби рівень глюкози був низьким.
- Були виділені мутанти кишкової палички, які не можуть бути спонукані метаболізувати будь-який цукор, крім глюкози. Існували дві загальні категорії мутантів:
- Клас I. Дефектний в ферменті аденілатциклази. Ці мутанти не в змозі зробити цАМФ навіть тоді, коли концентрація глюкози низька.
- Клас II. Не вистачає певного білка, відомого як білок рецепторів цАМФ (СРБ) або, також відомий як білок рецепторів катаболітів (СРБ).
- Максимальна транскрипція з лак-оперона вимагає наявності комплексу Camp/CRP.
- Комплекс Camp/CRP зв'язується з певною послідовністю в області контролю лаку, яка називається сайтом "CAP".
- Сайт CAP знаходиться лише вище за течією від місця зв'язування РНК полімерази.
- Мутації в місці CAP, що перешкоджають зв'язуванню Camp-CRP, також запобігають високому рівню експресії лак-оперону.
- Таким чином, зв'язаний комплекс Camp/CRP активує транскрипцію (позитивний контроль), тоді як зв'язаний лак-репресор пригнічує транскрипцію (негативний контроль).
- Комплекс Camp/CRP має спорідненість до ДНК та РНК pol.
- Підсилює комплексне утворення РНК pol з областю промотора ДНК.
Індукція лак-оперона аналогами лактози
- Лак-оперон може бути індукований лактозою
- b -галактозидаза (продукт гена lac Z) метаболізує лактозу
- Коли рівень лактози знижується, лак-оперон знову пригнічується лак-репресором (продукт гена лаку I)
- Неметаболізовані аналоги лактози можуть постійно індукувати (тобто де-репресувати) лак оперон
- ізопропіл б -тіогалактозид, або ІПТГ, є неметаболізованою аналогом лактози
Експерименти ДНК «слідів»
- Якщо білок зв'язується з областю ДНК, він може захистити цю область ДНК від перетравлення за допомогою dnase (dNaSe I: ендонуклеаза на ділянках, прилеглих до піримідинових нуклеотидів).
- Фрагмент ДНК може бути позначений на 5' закінчується 32 P, а потім мітка може бути переважно видалена з одного кінця (тобто 3' кінця гена) відповідною рестрикційної ендонуклеазою.
- Якщо цей фрагмент ДНК з міткою на одному конкретному кінці утворює комплекс з ДНК зв'язуючим білком білок, білок захистить область ДНК, з якою він зв'язується, від травлення dNaSe I.
- Травлення робиться так, щоб бути неповним, для цілей цієї дискусії уявіть, що кожна молекула ДНК розщеплюється лише один раз. Крім того, місце розщеплення вибирається випадковим чином з доступних ділянок.
- Фрагменти ДНК, відокремлені і проаналізовані за розміром (за допомогою гелевого електрофорезу) після травлення вказують на захищену область:
.png)
Малюнок 2.6.5: ДНК-сліди
Результати експериментів з друку слідів
Лак-ДНК інкубується або протеїном Camp/Cap, або РНК-полімеразою, або білком Lac I репресора:
.png)
Малюнок 2.6.6: Лак репресор з цАМФ
РНК-полімераза взаємодіє зі специфічними послідовностями промоторів і створює «слід» над областю ~ 70 пар основ.
- Цей захист спостерігався більш очевидним на одній пасмі, ніж на іншій (тобто якщо інша пасмо була позначена, результати не показали стільки захисту).
- Ця область захисту ДНКАзи включала ділянки в ДНК, з яких експерименти з мутагенезу виробляли або «вгору» регуляцію, або «вниз» регуляцію сили промотора.
- Ці мутагенні «гарячі» точки, що впливають на силу промотора, були розташовані на позиціях -10 або -30 вище за течією від місця початку транскрипції (позиція +1 на наведеній вище схемі):
.png)
Малюнок 2.6.7: Мутації сили промоутера
- Промоутери можна класифікувати за їх «силі».
- Це стосується відносної частоти ініціації транскрипції (події ініціації транскрипції в хвилину) і пов'язано з спорідненістю РНК-полімерази для промоторної області.
- Багато промоутерів в E. coli були охарактеризовані і була визначена послідовність промоутерів «консенсусу»:
.png)
Малюнок 2.6.8: Сила промоутера консенсусу
Примітка
Лак-промоутер є відносно слабким промоутером.
РНК-полімераза
- РНК-полімераза E. coli - це голофермент, що складається з субодиниць b ', b, a (димер) і s 70.
- Субодиниця s 70 - це субодиниця, яка зв'язується з промоторною областю, але не в змозі ініціювати синтез РНК.
- Після того, як субодиниця s 70 зв'язується, інші субодиниці зв'язуються, утворюючи функцію РНК-полімерази.
- Після того, як приблизно 10 пар основ були транскрибовані s 70 субодиниці листя і серцевина полімерази продовжується.
Регіон оператора Lac
- Область лак-оператора складається з (недосконалої) області перевернутого повторення.
- Не дивно, що активні молекули репресора складаються з гомодимера.
- У структурі гомодимера є пара областей a -спіралі, які вставляються в сусідні великі канавки ДНК.
- Поділ становить приблизно 34 ангстреми один від одного.