8.4: Розмноження ДНК у бактерій
- Page ID
- 6511

Доповнення
Джордж Бідл та Едвард Татум вперше описали концепцію того, що кожен ген відповідає ферменту в метаболічному шляху, піддаючи дріжджі Neurospora crassa мутагенним умовам (Beadle & Tatum, 1941). Після цих процедур Джошуа Ледерберг продовжив ці дослідження з Татумом, де вони створили два штами мутантів у кишковій паличці. Ці бактерії були ауксотрофами, не в змозі генерувати деякі основні поживні речовини, необхідні для підтримки їх росту. Два штами були описані як met − bio − Thr+ Leu+Thi + (Штам А) та Met + Bio + thr − leu − thi − (Штам B). Штам А може достатньо синтезувати амінокислоти треонін, лейцин і кофактор тіамін при дефіциті у виробництві кофактора біотину і амінокислоти метіоніну в той час як зворотне було вірно штаму B. Коли будь-який з цих двох штамів був нанесений на мінімальні середовища, зростання не відбулося. Доповнення мінімальних середовищ метіоніном і біотином дозволило штаму А рости в нормальному режимі. Коли два штами були змішані разом і покриті на мінімальних середовищах, спостерігався ріст бактерій. Два штами були здатні певним чином доповнювати один одного так, ніби стався статевий обмін генетичним матеріалом (Lederberg & Tatum, 1946).
Бактерії оснащені всіма необхідними ємностями для реплікації ДНК. Поширені види бактерій були адаптовані для використання в лабораторії для перенесення ДНК та розмноження її для використання в біотехнології. Крім хромосомної ДНК бактеріального генома, бактерії також мають екстрахромосомну ДНК, звану плазмідами. Ці плазміди реплікуються незалежно від бактеріальної хромосоми і можуть зустрічатися у високій копії. Ці кругові шматочки ДНК модифікуються в лабораторіях для перенесення конкретних шматочків ДНК, щоб їх можна було вивчати або використовувати для експресії в білки. Плазміди можуть природно нести важливі риси, включаючи стійкість до антибіотиків. Плазміди відносно невеликі, розміром від 1000 баз до 1 000 000 основ довжиною (1кб-1000kb).
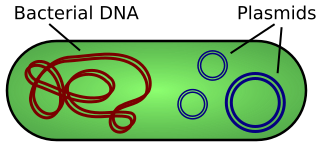
Бактеріальна ДНК зазвичай існує у вигляді великої кругової хромосоми (червоного кольору). Плазміди - це екстрахромосомні і автономно реплікуються шматочки ДНК (сині).
Через процес, який називається кон'югацією, бактерії можуть «статевим шляхом» передавати генетичний матеріал іншому, пропускаючи плазміди через структуру, звану кон'югаційним пілусом.

Процес кон'югації між донором, що несе плазміду, і реципієнтом без плазми. Донор створює кон'югаційний пілус для створення цитозольного моста з донором, де плазміда реплікується в реципієнт за допомогою методу реплікації кочення кола. Потім реципієнт стає компетентним виступати в якості донора.
особливості плазмід
Плазміди, розроблені біологами для човника шматочків ДНК для дослідження, називаються векторами, оскільки вони рухають шматок ДНК.

Ці плазмідні вектори мають ті ж ознаки, що і традиційні плазміди, здатні реплікуватися незалежно від бактеріального генома. Особливість, яка дозволяє реплікувати ці ДНК, називається походження реплікації (орі), яке зазвичай багате на А і Т, Однак ці плазмідні вектори мають додаткові властивості, які роблять їх легко працювати і відрізнити від бактеріальних плазмід; маркер виділення і багаторазовий сайт клонування. Маркер виділення зазвичай поставляється у вигляді гена, який кодує стійкість до конкретного антибіотика. У зображеній плазміді резистентність до ампіциліну надається геном β-лактамази. Сайт багаторазового клонування (MCS), також відомий як полілінкер, - це місце, в якому цікавить ДНК включена в вектор. MCSs визначаються набором унікальних ділянок, де ДНК може бути розрізана рестрикційними ендонуклеазами (РЕ). Як випливає з назви, рестрикційні ферменти «обмежені» в їх здатності різати або перетравлювати ДНК. Обмеження, яке корисно біологам, зазвичай є паліндромні послідовності ДНК. Паліндромні послідовності - це одна і та ж послідовність вперед і назад. Деякі приклади паліндромів: ГОНОЧНИЙ АВТОМОБІЛЬ, ГРОМАДЯНСЬКИЙ, ЛЮДИНА ПЛАН КАНАЛУ ПАНАМА. Що стосується ДНК, то є 2 нитки, які протікають протипаралельно один одному. Тому зворотне доповнення однієї пасма ідентично іншій.


eCori генерує липкі згуртовані кінці, SMAI генерує тупі кінці
Рестрикційні ферменти гідролізують ковалентні фосфодіефірні зв'язки ДНК, щоб залишити або «липкі/згуртовані» кінці, або «тупі» кінці. Ця відмінність у різанні важлива, оскільки липкий кінець eCori може бути використаний для узгодження шматочка ДНК, вирізаного з тим же ферментом, щоб склеїти або лігувати їх разом. Поки ендонуклеази розрізають ДНК, лігази з'єднують їх знову разом. ДНК, перетравлена за допомогою eCori, може бути перев'язана назад разом з іншим шматком ДНК, перетравленим за допомогою eCORI, але не до шматочка, перетравленого SMAI. Ще одним тупим різаком є eCorV з послідовністю розпізнавання GAT | ATC.
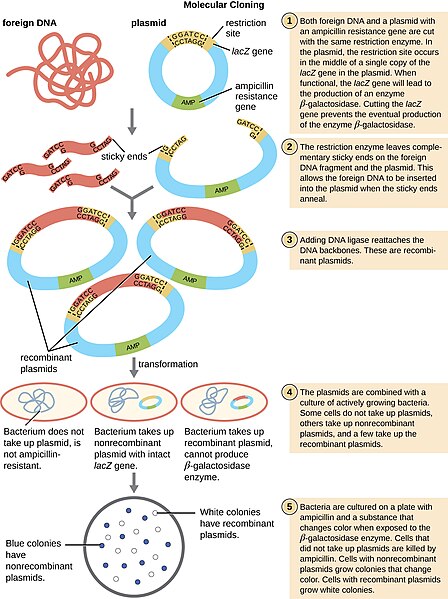
«розрізаючи і вставляючи» ДНК в вектори, ми можемо вводити чужорідні або екзогенні ДНК в бактерії. Цей тип ДНК зараз називається рекомбінантною ДНК і є серцем біотехнології.
Технологія рекомбінантної ДНК

Додаткові ресурси
Питання для роздумів
1. Чому ви думаєте, що витоки реплікації складаються з А і Т?
2. Чим відрізняються типи зв'язків, що тримають подвійні нитки разом, проти фосфодіефірних зв'язків хребта ДНК?
3. Чи можна перетравлювати ДНК за допомогою SMAI, перев'язати до ДНК, перетравленої за допомогою eCorV?
4. Якщо так, то який фермент зможе перетравлювати цю нову ДНК?
- Бідл, Г.В.; Татум, Е.Л. (1941). «Генетичний контроль біохімічних реакцій в нейроспорі». Збірник наукових праць Національної академії наук. 27 (11): 499-506. дої: 10,173/п.27.11.499. ПМК 1078370. ІПМІД 16588492
- Ледерберг Дж., Татум Е.Л. «Рекомбінація генів в кишковій паличці». Природа. 158 (4016): 558. Код: 10.1038/15855a0