6.2: Птеридофіта - папороті
- Page ID
- 6582

Птеридофіта, папороті і союзники мають приблизно 12 000 видів і шість класів (рис.\(\PageIndex{1}\)). Вони мають споричний життєвий цикл із переважанням спорофітів, тоді як їх гаметофіти часто зводяться до проталію, невеликої рогоподібної рослини. Ще один частий варіант - підземний, мікопаразитарний гаметофіт. Птеридофіти (за одним винятком) мають справжні коріння. Більшість з них мають судинні тканини і є гомогідрогенними. Саме тому насіннєві рослини разом з папоротями мають назву судинні рослини. Pteridophyta sporophytes завжди починають своє життя з ембріона, розташованого на гаметофіті. Хоча у Pteridophyta є справжня ксилема і флоема, вони не мають розвиненого вторинного потовщення.
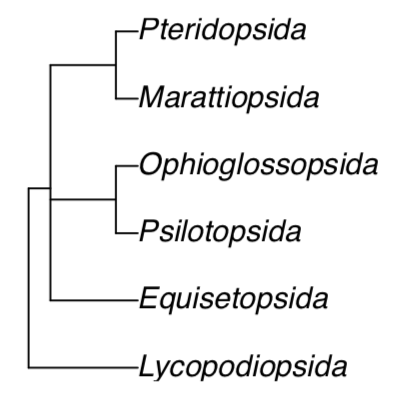
Найдавніші птеридофіти з'явилися в силурійський період, вони були риніофітами. Риніофіли мали добре розвинені надземні гаметофіти і відносно короткі, дихотомно розгалужені безлисті спорофіти. Наступними важливими етапами стали формування листя і подальше скорочення гаметофітів.
різноманітність птерідофітів
Lycopodiopsida, або лікофіти мають щонайменше чотири пологи і більше 1200 видів. Лікофіти належать до мікрофільозного роду птерідогітів. Це означає, що їх листя виникли від появи поверхні стебла, а тому більше схожі на листя моху, ніж будь-які інші листя птеридофітів і насіннєвих рослин. Lycophyte sporangia пов'язані з листям і часто утворюють стробілус, який є конденсацією спорангійних листя (спорофілів, коли вони листоподібні або спорангіофори, коли вони розходяться). Їх сперматозоїди зазвичай мають 2 джгутика (як мохи), але іноді також мультиджгутикові (як сперматозоїди інших папоротей). Лікофіти раніше були домінуючими рослинами кам'яновугільних тропічних болотних лісів, а їх залишки стали вугіллям. Сучасні лікофіти набагато менші, але все ще процвітають у вологих і теплих місцях. Більше базальних лікофітів (клубних мохів Huperzia та Lycopodium) мають рівні спори та підземні гаметофіти, тоді як більш просунуті Selaginella (spikemoss) та Isoëtes (перо) обидва гетероспористі (див. Нижче) зі зменшеними надземними гаметофітами. Квілворт є прямим нащадком гігантських вугільних лікофітних дерев, і, незважаючи на те, що є підводним гідрофітом, він все ще зберігає незвичайне вторинне потовщення стебла. Багато колосових мохів є пойкілогідричними (ще одна схожість з мохами).
Equisetopsida (хвощі) - це невелика група з одним родом, Equisetum, і налічує близько 30 різних трав'янистих видів, які зазвичай живуть у вологих місцях проживання. Листя цих рослин зведені в лусочки, а стебла сегментовані і також фотосинтезируются; є і підземне кореневище. Стебловий епідерміс містить кремнезем, що робить його абразивною поверхнею, і через це американські піонери використовували б цю рослину для очищення каструль і сковорідок. Саме так воно і отримало прізвисько «чистячий пік». Стебло має множинні канали, це якось схоже на стебла трав. Спорангії пов'язані з шестикутними стеблими спорангіофорами; є також елатери, які є не окремими клітинами, а частинами спорової стінки. Гаметофіти, як правило, дрібні і дводомні, але самі рослини гомоспорні: менші пригнічені гаметофіти розвивають лише антеридії, тоді як більші гаметофіти розвивають лише архегонію.
Псилотопсіда (віночкові папороті) - це невелика тропічна група, яка складається лише з двох пологів, Psilotum і Tmesipteris, що налічує лише сім різних видів. Це трав'янисті рослини, які ростуть як епіфіти. Папороті віночків гомоспористі, а їх спорангії зливаються в синангию adnate sporangia. Псилотопсіди мають протостели, як деякі лікофіти, і довгоживучі підземні гаметофіти; у них також є багатобічеподібні сперматозоїди, схожі на всі інші папороті. І Psilotum, і Tmesipteris не мають коренів; крім того, Psilotum також не вистачає листя.
Ophioglossopsida (язикові папороті) - невелика група, яка складається приблизно з 75 видів, і є найближчими родичами для віночок папоротей. Офіоглоссопсида має підземне кореневище (іноді зі слідами вторинного загущення) з надземними двосекційними листям: одна половина кожного листа - листова пластинка, а інша половина стає спорофілом. Гаметофіти також ростуть під землею. Ophioglossum vulgatum, відомий також як папороть язика гадюки, має хромосомне число\(2n=\) 1,360, що є найбільшим хромосомним числом коли-небудь!
Мараттіопсіда (гігантські папороті) - тропічні рослини, з декількома пологами і близько 100 видів. Вони схожі на справжні папороті і мають складні листя, які згортаються в молодості. Вони також є найбільшими папоротями, оскільки один лист може становити шість метрів в довжину. У них короткі стебла, а листя з прилистниками. Їх спорангії мають багатошарові стінки і зливаються в синангії (не схожі на справжні папороті). При цьому вони розташовуються на нижній поверхні листя (як у справжніх папоротей). Гаметофіти відносно великі (1—2 см), фотосинтетичні, і зазвичай живуть тривалий час. Ці папороті були важливі в кам'яновугільних болотних лісах.
Pteridopsida (справжні папороті) налічують понад 10 000 видів і становлять більшість живих монолофітів (всі класи Pteridophyta, крім лікофітів). Їх листя називають листям через верхівкового зростання; молоді листя згортаються в скрипки (рис.\(\PageIndex{2}\)). Справжні папороті мегафілові: їх листя походять від сплюснутих гілок. Справжні папороті мають унікальні спорангії: лептоспорангія. Лептоспорангії походять з однієї клітини в листі, у них довгі тонкі стебла, і стінка одного клітинного шару; вони також активно відкриваються: коли спорангіум дозріє (висихає), ряд клітин з потовщеними стінками (кільцевого кільця) буде скорочуватися повільніше навколишніх клітин і нарешті зламається і звільняється всі суперечки відразу. Лептоспорангії також групуються в скупчення, які називаються сорі, які часто покриті парасолькою або кишеньковою індузією. Гаметофіти Птеридопсида бувають дрібними і ростуть надземними. Деякі пологи справжніх папоротей (наприклад, комариний папороть Azolla, водяний трилисник Marsilea і кілька інших) є гетероспорими.

Справжні папороті відрізняються високою конкуренцією навіть покритонасінним рослинам. Незважаючи на свій «примітивний» життєвий цикл, вони мають безліч переваг: здатність фотосинтезуватися в глибокій тіні (вони не зобов'язані швидко рости), виживати підвищену вологість, заробляти мільярди репродуктивних одиниць (спор). Папороті не потрібно витрачати свої ресурси на квіти та плоди, а також менш вразливі до хребетних травоїдних тварин та комах-шкідників, ймовірно, тому, що вони не використовують їх як запилювачів і, отже, можуть отруювати тканини проти всіх тварин.
Гетероспори: Наступний крок на суші
Хребетні тварини стали повністю наземними (земноводні стали першими рептиліями) тільки тоді, коли їх запліднення стало повністю незалежним від води. Подібні «еволюційні зусилля» рослини почали виконувати ще раніше, але поки плазуни активно наближаються до статевого партнера, рослини не можуть зробити те ж саме, оскільки їх тканини і органи розвивалися для абсолютно різних цілей. Замість активної статі рослини використовують «килимове бомбардування» спорами; це було винайдено, щоб збільшити ймовірність того, що дві суперечки приземляться поруч і відстань між сперматозоїдом і яйцеклітиною буде мінімальним.
Однак, оскільки просте збільшення кількості суперечок є великою марною тратою ресурсів, рослини мінімізують розміри спор; це також дозволить збільшити відстань розгону. З іншого боку, деякі спори повинні залишатися великими, оскільки ембріон (якщо відбудеться запліднення) потребуватиме підтримки від годування гаметофіту. Отже, рослини закінчилися поділом праці: численні, мізерні чоловічі спори, які переростають у чоловічих гаметофітів лише з антеридіями, і мало великих жіночих суперечок, які роблять жіночих гаметофітів, що виробляють лише архегонію (рис.\(\PageIndex{3}\)).

Цей гетероспоричний цикл робить запліднення менш залежним від води і більш залежним від розподілу спор і особливостей гаметофіту (рис.\(\PageIndex{4}\)). Це також дозволяє численні вдосконалення в майбутньому.
Поділ праці дозволяє використовувати ресурси ефективніше, а також обмежує самозапліднення. В еволюції рослин була висока потреба в гетероспорії, оскільки вона самостійно виникла в декількох групах птерідофітів і навіть серед мохів. В крайніх випадках гетероспори (рис.\(\PageIndex{5}\)) жіноча спора не залишає материнську рослину і проростає там, «чекаючи» запліднення від розвиненого поблизу самця гаметофіта; по суті, це початкове запилення, крок до насіння.
Гетероспорові рослини виробляють одну жіночу спору, мегаспору, яка багата поживними речовинами; мегаспори не широко розпорошені, але що виходить з неї самка гаметофіта забезпечує живлення і захист зиготи, ембріона та молодого спорофіта. Гетероспоричний життєвий цикл (рис.\(\PageIndex{6}\)) починається з чоловічого гаметофіта і жіночого гаметофіта, обидва з яких виробляють гамети. Як тільки відбувається запліднення, зигота переростає в спорофіт. Потім спорофіт вироблятиме два різних типи спорангії: жіноча мегаспорангія та чоловіча мікросоргангія. Мейоз у мегаспорангіумі часто призведе до однієї жіночої спори, мегаспор (подібно до мейозу в яєчниках хребетних тварин), тоді як в мікроспорангії мейоз та наступні мітози зроблять численні мікроспори; як мегаспор, так і мікроспори переростуть в гаметофіти і цикл повториться.

Загалом, гетероспорія дозволяє розділити між чоловічими та жіночими гаплоїдними лініями. Самці гаметофіти стають настільки маленькими, що їх можна було легко транспортувати в цілому. Цілі чоловічі гаметофіти починають бути рухомою стадією - це походження запилення.

