8.2: Синдроми запилення
- Page ID
- 7026

Цілі навчання
- Поясніть стратегії запилення щодо генетичного різноманіття.
- Поясніть, що мається на увазі під терміном синдром запилення.
- Використовуйте квіткові характеристики, щоб передбачити рослин-запилювач (и).
У покритонасінних рослин запилення визначається як розміщення або перенесення пилку з пильовика на рильце того ж квітки або іншої квітки. У голонасінних рослин запилення передбачає перенесення пилку з чоловічої шишки в жіночу шишку. Після перенесення пилок проростає, утворюючи пилкову трубку та сперму для запліднення яйцеклітини.
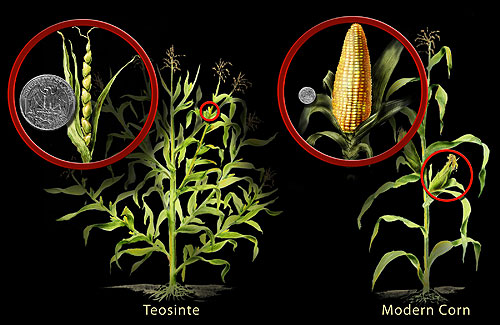
Запилення добре вивчено ще з часів Грегора Менделя. Мендель успішно здійснював самозапилення, а також перехресне запилення гороху, вивчаючи, як передавалися характеристики з одного покоління в інше. Сьогоднішні культури є результатом селекції рослин, яка використовує штучний відбір для отримання сучасних сортів (термін, який відноситься до культурних сортів). Прикладом є сьогоднішня кукурудза, яка є результатом років селекції, розпочатої з її предка, трави під назвою теосінт (див. Рисунок\(\PageIndex{1}\)). Теосінт, який спочатку почали культивувати стародавні майя, мав невеликі насіння - значно відрізняються від сьогоднішніх відносно гігантських колосків кукурудзи та пухких ядер. Цікаво, що хоча ці дві рослини здаються абсолютно різними, генетична різниця між ними незначна.
.jpg)
.jpg)
Запилення та генетичне різноманіття
Запилення приймає дві форми: самозапилення і перехресне запилення. Самозапилення відбувається, коли пилок з пильовика відкладається на рильце того ж квітки, або іншої квітки на тому ж рослині. Самозапилення відбувається в квітках, де тичинка і килима дозрівають одночасно, і розташовані так, що пилок може приземлитися на рильце квітки. Цей метод запилення не вимагає інвестицій від рослини для забезпечення нектару та пилку в якості їжі для запилювачів, але призводить до зниження генетичної різноманітності в популяції. Самозапилення часто існує як «план Б», в разі, якщо перехресне запилення з якихось причин неможливо. Іноді самозапильні квітки навіть не розкриваються, ці квіти називають клейстогамними.
Перехресне запилення - це перенесення пилку з пильовика однієї квітки на рильце іншої квітки на різних особин одного виду. Цей метод спирається на вектор пилку, але призводить до більш високого генетичного різноманіття.
Живі види пристосувалися для забезпечення виживання свого потомства; ті, які не вдаються, вимерли. Тому генетичне різноманіття потрібно, щоб при зміні умов навколишнього середовища або стресу частина потомства могла вижити. Самозапилення призводить до виробництва рослин з меншим генетичним різноманіттям, так як генетичний матеріал з одного і того ж рослини використовується для формування гамет, а в кінцевому підсумку і зиготи. На відміну від цього, перехресне запилення - або перехрещення - призводить до більшої генетичної різноманітності, оскільки мікрогаметофіт і мегагаметофіт походять від різних рослин.
Оскільки перехресне запилення дозволяє отримати більше генетичного різноманіття, рослини розробили багато способів уникнути самозапилення. У деяких видів пилок і зав'язі дозрівають в різний час. Ці квіти роблять самозапилення практично неможливим. До моменту дозрівання і скидання пилку рильце цієї квітки дозріває і запилюватися може тільки пилком з іншої квітки. Деякі квіти мають розвинені фізичні особливості, що перешкоджають самозапиленню. Примула - один з таких квіток. Первоцвіти розвинули два типи квітів з відмінностями в довжині пильовика та рильця: квітка з рожевими очима має пильовики, розташовані на півдорозі пилкової трубки, а рильце квітки молочниці також розташоване на півдорозі. Комахи легко перехресно запилюються, шукаючи нектар на дні пилкової трубки. Це явище також відоме як гетеростилія. Багато рослин, такі як огірок, мають чоловічі та жіночі квіти, розташовані на різних ділянках рослини, що ускладнює самозапилення. У ще інших видів чоловічі та жіночі квіти виносяться на різних рослинях (дводомних). Все це є бар'єрами для самозапилення, тому рослини залежать від запилювачів для перенесення пилку. Більшість запилювачів є біотичними агентами, такими як комахи (наприклад, бджоли, мухи та метелики), кажани, птахи та інші тварини. Інші види рослин запилюються абіотичними агентами, такими як вітер і вода.
Підсумовуючи, самонесумісність - це механізм, який перешкоджає самозаплідненню багатьох видів квітучих рослин для збільшення генетичного різноманіття на рівні популяції. Робота цього механізму самонесумісності має важливі наслідки для селекціонерів, оскільки гальмує виробництво інбредних та гібридних рослин.
Гени несумісності в квітах
В останні десятиліття гени несумісності - які перешкоджають проростанню пилку або переростанню в стигму квітки - були виявлені у багатьох видів покритосперми. Якщо рослини не мають сумісних генів, пилкова трубка припиняє зростання. Самонесумісність контролюється локусом S (стерильність). Пилкові трубки повинні рости через тканину рильця і стиль, перш ніж вони зможуть потрапити в яйцеклітину. Карпель селективний за типом пилку, він дозволяє рости всередині. Взаємодія відбувається насамперед між пилком і клітинами епідермісу стигми. У деяких рослин, таких як капуста, пилок відторгається на поверхні рильця, а небажана пилок не проростає. У інших рослин проростання пилкової трубки затримується після зростання на третину довжини стилю, що призводить до загибелі пилкової трубки. Загибель пилкової трубки обумовлена або апоптозом (запрограмованою загибеллю клітин), або деградацією РНК пилкової трубки. Деградація є результатом активності рибонуклеази, кодованої локусом S. Рибонуклеаза виділяється з клітин стилю в позаклітинному матриксі, який лежить поряд з зростаючою пилкової трубкою.
Синдроми запилення
Згодом покритонасінні рослини еволюціонували різні морфології, запахи та кольори квітів, які відповідали їх конкретному вектору пилку. Ці набори характеристик, звані синдромами запилення, дозволяють вченим прогнозувати запилювачі для різних рослин. Наприклад, чашоподібні квіти зазвичай запилюються масивними тваринами, такими як жуки і навіть кажани. Лійкоподібні квіти, а також губі квіти (з губами), пристосовані для мух і бджіл. Квіти з довгими шпорами приваблюють метеликів і птахів (на зразок колібрі або цукрових птахів). Так само багато організмальних векторів розвивали особливості або поведінку, щоб спеціалізуватися на запиленні певних рослин. Ця еволюція тандему до збільшення специфічності називається коеволюцією. Цікаво, що фенотипи квітів також можуть співвідноситися з іншими факторами навколишнього середовища (наприклад, Peach et al. 2020). Додаткові зображення синдромів запилення див. Розділ 14.1 Синдроми запилення у Фотографічному атласі.
Запилення обманом
Орхідеї високо цінуються квітами, з безліччю рідкісних сортів (рис.\(\PageIndex{2}\)). Вони ростуть в діапазоні специфічних середовищ існування, переважно в тропіках Азії, Південної Америки та Центральної Америки. Виявлено щонайменше 25 000 видів орхідей.

Квіти часто залучають запилювачів харчовими нагородами, у вигляді нектару. Однак деякі види орхідей є винятком з цього стандарту: вони еволюціонували різні способи залучення бажаних запилювачів. Вони використовують метод, відомий як харчовий обман, при якому пропонуються яскраві кольори та парфуми, але не їжа. Anacamptis morio, широко відомий як зелено-крила орхідея, несе яскраво-фіолетові квіти і випромінює сильний аромат. Джміль, його головний запилювач, приваблює квітка через сильний запах, який зазвичай вказує на їжу для бджоли - і в процесі підбирає пилок для транспортування до іншої квітки.
Інші орхідеї використовують сексуальний обман. Chiloglottis trapeziformis випромінює з'єднання, яке пахне так само, як феромон, що виділяється самкою оси для залучення чоловічих ос. Самець оса приваблює запах, приземляється на квітку орхідеї, і в процесі переносить пилок. Деякі орхідеї, як австралійська орхідея молотка, використовують аромат, а також візуальну хитрість у ще одній стратегії сексуального обману для залучення ос. Квітка цієї орхідеї імітує зовнішній вигляд жіночої оси і виділяє феромон. Самець оса намагається спаровуватися з тим, що здається самкою оси, і в процесі підбирає пилок, яку потім передає наступному підробленому мату.
Бджоли
Бджоли, мабуть, найважливіший запилювач багатьох садових рослин і більшості комерційних плодових дерев (рис.\(\PageIndex{3}\)). Більшість бджіл, яких люди помічають, - це соціальні бджоли, джмелі та медоносні бджоли, хоча існують тисячі видів одиночних бджіл, які мають важливу роль запилення. Бджоли мають розгалужені волоски, що покривають їх тіла, що робить їх чудовими запилювачами, оскільки більше пилкових зерен, ймовірно, потраплять на їх зовнішні поверхні та переносяться на інші рослини.
Бджоли збирають багаті енергією пилок і нектар для своїх потреб у виживанні та енергії, тому квіти, які спільно розвивалися з бджолами, містять як нектар, так і пилок. Оскільки бджоли не бачать червоного кольору, бджолозапильні квіти зазвичай мають відтінки синього, жовтого, фіолетового або іншого кольору. Вони відвідують квіти, які відкриті протягом дня, яскраво забарвлені, мають сильний аромат або аромат, з зоною посадки якогось виду, як правило, з наявністю нектарового направляючого. Посібник з нектару може включати регіони на квітці з пігментами, які видно лише бджолам, а не людям (наприклад, УФ); це допомагає направляти бджіл до центру квітки, тим самим роблячи процес запилення більш ефективним. Пилок прилипає до нечіткої шерсті бджіл, а коли бджола відвідує іншу квітку, частина пилку переноситься на другу квітку.


Останнім часом популяції медоносних бджіл (інтродукованого європейського виду) знижуються через безліч факторів, включаючи пестициди (наприклад, неонікотиноїди) та паразитичних кліщів. Оскільки одиночні бджоли не експлуатуються комерційною промисловістю, вплив на ці популяції був менш добре задокументований. Однак зниження популяції бджіл матиме надзвичайно важливий вплив на нашу продовольчу безпеку, оскільки бджоли використовуються для запилення багатьох харчових культур.
Оси також є важливими комахами-запилювачами, і запилюють багато видів інжиру.
мухи
Багато видів мух є важливими запилювачами. Ці мухи часто виглядають і поводяться так само, як бджоли, виконуючи важливі ролі в запиленні. Однак, оскільки ці квіти, як правило, класифікуються під синдром запилення бджіл, ми натомість обговоримо другу категорію мух для синдрому запилення мух. Ці комахи (мухи, а іноді і жуки-падлі) зазвичай шукають не квітку, а труп або якийсь гній, щоб харчуватися або відкладати яйце. Деякі квіти еволюціонували, щоб обманювати мух до запилення, випромінюючи гниючий запах або запах гниючої плоті. Ці квіти, які виробляють нектар, зазвичай мають тьмяні кольори, такі як коричневий або фіолетовий. Вони часто мають якусь функцію, подібну до пастки, тому, коли муха відвідує, вона повинна трохи підстрибувати всередині квітки перед від'їздом (це збільшує ймовірність відкладення пилку, як на квітці, так і на льоту). Приклади включають квіти трупа (Amorphophallus і Rafflesia) і дракон Арум (Dracunculus).

Метелики і метелики
Метелики, такі як монарх, запилюють багато садових квітів і польових квітів, які зазвичай зустрічаються гронами. Ці квіти яскраво забарвлені, мають сильний аромат, відкриті протягом дня і мають направляючі нектару, щоб полегшити доступ до нектару. Пилок підбирають і переносять на кінцівках метелика.
З іншого боку, молі запилюють квіти пізно вдень і вночі. Квітки, запилені моллю, бліді або білі і плоскі, що дозволяє молі приземлитися. Одним з добре вивчених прикладів рослини, що запилюється міллю, є рослина юка, яка запилюється міллю юки. Форма квітки і молі пристосувалися таким чином, щоб забезпечити успішне запилення. Моль відкладає пилок на липкому рильце, щоб запліднення відбулося пізніше. Самка молі також відкладає яйцеклітини в яєчник. Оскільки яйцеклітини розвиваються в личинок, вони отримують їжу з квітки і розвиваються насіння. Таким чином, і комаха, і квітка виграють один від одного в цьому симбіотичному відношенні. Аналогічне співвідношення мають кукурудзяна вушна моль і рослина Гаура (рис.\(\PageIndex{5}\)).

кажани
У тропіках і пустель кажани часто є запилювачами нічних квітів, таких як агава, гуава та іпомея. Квітки, як правило, великі і білі або блідо-забарвлені; таким чином, їх можна відрізнити від темного оточення вночі. Квіти мають сильний, фруктовий або мускусний аромат і виробляють велику кількість нектару. Вони від природи великі і з широким ротом для розміщення голови кажана (рис.\(\PageIndex{6}\)). Коли кажани шукають нектар, їх обличчя і голови покриваються пилком, яка потім переноситься на наступну квітку.

Птахи
Багато видів дрібних птахів, такі як колібрі (рис.\(\PageIndex{7}\)) і сонячні птахи, є запилювачами для таких рослин, як орхідеї та інші польові квіти. Квіти, які відвідують птахи, зазвичай міцні і орієнтовані таким чином, щоб дозволити птахам залишатися біля квітки, не заплутавшись крила в сусідніх квітах, таких як бовтаються (кулон). Квітка зазвичай має вигнуту трубчасту форму, що забезпечує доступ для дзьоба птаха. Яскраво забарвлені квітки без запаху, які відкриваються протягом дня, запилюються птахами. Коли птах шукає багатий енергією нектар, пилок осідає на голові та шиї птиці, а потім переноситься до наступної квітки, яку вона відвідує. Відомо, що ботаніки визначають ареал вимерлих рослин шляхом збору та ідентифікації пилку з 200-річних зразків птахів з тієї ж ділянки.

Вітер
Більшість видів хвойних дерев, а також багато покритонасінні, такі як трави, клени і дуби, запилюються вітром. Соснові шишки коричневі і без запаху, тоді як квіти вітрозапилюваних покритонасінних видів зазвичай зелені, дрібні, можуть мати маленькі пелюстки або не мати, і виробляють велику кількість пилку. На відміну від типових квіток, що запилюються комахами, квіти, пристосовані до запилення вітром, не виробляють нектару або запаху. У вітрозапилюваних покритонасінних видів пильовики часто великі і звисають з квітки, і, як дме вітер, легку пилок переносять з собою (рис.\(\PageIndex{8}\)). Квітки зазвичай з'являються рано навесні, перед листям, щоб листя не блокували рух вітру. Пилок відкладається на оголеному пір'ястому рильце квітки (рис.\(\PageIndex{9}\)).


Вода
Деякі рослини, такі як австралійська морська трава і ставкові бур'яни, запилюються водою. Пилок плаває на воді, і при зіткненні з квіткою відкладається всередині квітки.
Зведена таблиця синдромів запилення
Таблиця\(\PageIndex{1}\) підсумовує синдроми запилення, описані в категоріях вище.
| Синдром запилення | Колір | Структура | Аромат | Нектар або пилок |
|---|---|---|---|---|
| Вітер | Тупий, оцвітина часто відсутня або зменшена | Великі пір'ясті рильця, великі пильовики | Жоден | Без нектару, великої кількості пилку |
| Птахи | Червоні і рожеві | Часто трубчасті або чашечкові | Жоден | Багато прихованого нектару, помірний пилок |
| Бджоли | Фіолетовий, синій, жовтий, білий, УФ | Плоскі і неглибокі або трубчасті, з майданчиком посадки | Солодкий, свіжий, м'який | Пилок часто липкий і запашний, нектар зазвичай присутній |
| кажани | Білий, тьмяно-зелений або фіолетовий | Часто чашеподібні або підвісні, пильовики стирчать | Затхлий або фруктовий, сильний, що випускається вночі | Багато прихованого нектару |
| Метелики | Білий, блідо-рожевий або фіолетовий | Часто трубчасті або чашечки, без посадкової площадки | Міцний і солодкий, що випускається вночі | Багато прихованого нектару, обмежена пилок |
| Метелики | Яскраві кольори | Трубчасті, з широкою посадковою площадкою | Слабкий, свіжий | Багато прихованого нектару, обмежена пилок |
| мухи | Темно-червоний, фіолетовий, коричневий | Неглибокий, воронковий або пастковий | Гнильний, гниючий | Без нектару, помірного пилку |
Атрибуції
Куратор і автор Марія Морроу з використанням наступних джерел:
- 32.2 Запилення та запліднення з біології 2e OpenStax (ліцензований CC-BY). Доступ безкоштовно на openstax.org.
- 8.2 Квітка і плід з вступу в ботаніку Олексія Шипунова (суспільне надбання)